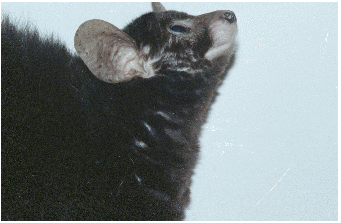
Abbildung 3.1: Heinrich Sprankel in Salem
Die Umstände des Kennenlernens waren unterschiedlich. Als Anekdote möchte ich hier
von Ludwig Wolf berichten. Als Wolf sich um das Amt des Kanzlers in Gießen bewarb,
war ich der stellvertretende Vorsitzende des AStAs der Universität und studentisches
Mitglied im Kleinen Senat, dem damals höchsten Gremium der Universität. Vom
ASTA der Universität Mainz wurde ich informiert, dass Wolf CDU-Mitglied sei, dies
wollten wir bei der Senatsanhörung „herausarbeiten“. Gleich den ersten Bewerber,
einem Mitglied der SPD, fragte ich, ob er Mitglied einer politischen Partei sei. Der
Vorsitzende ließ diese Frage nicht zu, und nach der Anhörung wurde ich gerügt, so
eine Frage stelle man nicht. Als sich dann Wolf vorstellte, fragte ich ihn konkret,
ob er Mitglied der NPD sei, diese Frage konnte man mir nicht verbieten.
Erwartungsgemäß war die Antwort ein „Nein“. Gleichzeitig hatte ich aber
unbeabsichtigt auf mich aufmerksam gemacht. Es war nämlich die Frage, die Wolf –
zurück in Mainz – seiner Frau berichtete: „Stell dir vor, man hat mich gefragt, ob ich
NPD – Mitglied sei“.
Monate später, im Gespräch mit meinem Freund Dr. Jürgen H. Jungbluth im
Tierhaus der Universität hatte ich nebenbei erfahren, dass ein neuer Professor
(Sprankel) gerne Primaten aus dem MPI für Hirnforschung in Frankfurt mitbringen
wolle. Von der Anthropologie, die einen Affenraum (ohne Affen) im Tierhaus hatte,
wusste ich, dass Professor Schäfer auch gerne lebende Primaten halten würde. Ich
beschloss, die Interessen beider zu bündeln, ging zu Schäfer und sagte, ich könnte
Primaten bekommen, ich bräuchte nur einen Raum, und ging zu Sprankel und sagte,
ich würde gerne meine Diplomarbeit über Primaten schreiben, ich hätte die
Möglichkeit, einen Raum zu bekommen, bräuchte aber noch Primaten. Beide Herren
waren sehr interessiert, nur dürfte es nichts kosten, beide hatten kein Geld. Also ging
ich zum Kanzler der Universität, nun Ludwig Wolf, der sich gleich an mich erinnerte,
und fragte ihn um Rat, ich hätte Räumlichkeiten, Primaten, aber kein Geld, um sie zu
ernähren. Wolf fragte nach der Summe, DM 1500 pro Jahr, und sagte „Kein
Problem“, ich bekäme einen eigenen Etat, Sprankel müsste die Rechnungen nur
unterschreiben.
Am 03. Februar 1971 war es endlich soweit. Ich erinnere mich nur, dass ich die vier
Riesengalagos in Frankfurt, im MPI für Hirnforschung, abgeholt habe, wie weiß ich
nicht mehr, wahrscheinlich mit Hilfe der Fahrbereitschaft der Universität.
Von dort erhielt ich auch ihre Schlafkästen geborgt und wurde von dem Tierpfleger
Günter Oberländer mit der Diät vertraut gemacht. So begann meine Karriere
als Primatologe. Von Sprankel erhielt ich das Thema: „Duftmarkieren beim
Riesengalago“ und die Negativa einer abfotographierten Arbeit von Eibl-Eibesfeldt
([46]). Zurück in Gießen kümmerte ich mich um meine Versuchstiere und bat
den Direktor des I. Zoologischen Instituts, Prof. Dr. Armin Wessing, um die
Gefälligkeit, seiner Fotolaborantin zu gestatten, für mich die vier Seiten der
Arbeit von Eibl-Eibesfeldt zu entwickeln, was Wessing zuließ. Er erlaubte
mir auch, dem hauseigenen Schreiner Aufträge zu erteilen (wenn es nichts
kostet).
Da alle Tiere einen lateinischen Namen haben, musste ich diesen zuerst erkunden.
Dies war schwieriger als gedacht und zog sich über viele Jahre hin. Da ich in diesem
Buch nicht mehrfach den Namen meiner Tiere ändern möchte, sei als erstes über
diesen Teil meiner langjährigen Arbeit berichtet.
Der erste Riesengalago wurde 1812 von E. Geoffroy beschrieben, heißt also korrekt Galago crassicaudatus
E. Geoffroy, 1812.1
In der damals modernsten Darstellung der Primaten gab der bekannte W. C. Osman Hill
( [85]) auch einen Bestimmungsschlüssel an, nach diesem handelte es sich bei meinen
Tieren wohl um Galago crassicaudatus kikuyuensis. Ich lernte auch, dass es verschiedene
Unterarten des Riesengalagos gebe. Hill verschwieg auch nicht, dass diskutiert worden
sei, zwischen nördlichen kleineren und südlichen größeren Unterarten auf Artenniveau
zu differenzieren. Hill folgte aber dem deutschen Bearbeiter der Riesengalagos Schwarz
([198]) und stellte in Abrede, dass es verschiedene Arten der Riesengalagos
gebe.2
Im Prinzip übernahm Hill kritiklos jedoch die Einschätzung von Schwarz über die verschiedenen
Riesengalago-Unterarten3.
Dieser sind dann alle weiteren Galago Bearbeiter gefolgt. Sehr einflussreich für
die „korrekte“ Bestimmung der Unterart war dann eine Arbeit von John
Buettner-Janusch ([13]), der ausführte, es gebe bei den Riesengalgos nur zwei
Unterarten, nämlich Galago crassicaudatus crassicaudatus und Galago crassicaudatus
argentatus.

Danach waren meine Tiere „eindeutig“ Galago crassicaudatus crassicaudatus, so habe ich auch deren Namen in der Diplomarbeit angegeben. Meine Annahme, dass Buettner-Janusch korrekt lag mit seiner Einschätzung, wurde fragwürdig, als ich im November 1973 von Prof. H. K. Dennig in München ein Paar Riesengalagos erhielt, die anders aussahen als unsere Tiere.
Wir haben sie als Galago crassicaudatus panganiensis identifiziert. Am 22. April 1976 – bereits in Kassel – erhielt ich dann ein Paar Galago crassicaudatus argentatus4, die nicht nur völlig anders aussahen sondern sich auch völlig anders verhielten als unsere Tiere. Sie waren mit Sicherheit eine andere Art.5 Aber nur eine konnte Galago crassicaudatus sein. Daher fuhren meine Frau und ich 1979 nach Paris, um Geoffroys Galago zu besuchen. Dieser war in einem schlechten Zustand, doch hatte ich nach der Größe des Schädels den Eindruck, unsere argentatus müssten crassicaudatus sein. Insofern müssten unsere ursprünglichen Galagos einen neuen Artnamen erhalten. Dieser war schnell zu finden. Der älteste Name der nördlichen Galagos war Galago agisymbanus Coquerel, 1859. Insofern mussten unsere Galagos nun den Artnamen Galago agisymbanus führen. Diese eigentlich richtige Einschätzung erwies sich aber als vorläufig. Todd Olson6 hatte in einer bewundernswerten Neubearbeitung der Riesengalagos ([150]), die leider erst 1982 publiziert wurde, herausgefunden, dass der nach Hill und Schwarz südlichste Vertreter der Riesengalagos, der 1838 durch Ogilby beschriebene Garnett-Galago, überhaupt kein südlicher Vertreter war. Wie in einem Krimi entwirrte Olson ([150]), wie dieser 1862 den falschen Fundort Port Natal erhielt.7 Tatsächlich wäre der beschriebene Galago ein Vertreter der nördlichen Unterarten. Der Name des 1838 beschriebenen Galagos hat Priorität vor dem Namen des von Coquerel erst 1859 beschriebenen Individuums. Unsere Tiere sind demnach Galago garnettii, konkret Galago garnettii garnettii Ogilby, 1838 (vgl. auch Welker und Schäfer–Witt [282])8.
Nach dem Bestimmen der Riesengalagos begann die Literatursuche. Diese war damals – ohne Internet – schwierig. Ich durchsuchte systematisch alle einschlägigen Zeitschriftenbände und versuchte über die Fernleihe die Quellen zu erhalten, die unterschiedliche Autoren genutzt hatten, ging – angefangen mit der schon erwähnten Arbeit von Eibl-Eibesfeldt ([46]) – Zitat für Zitat durch und stellte schnell fest, dass einige Autoren nahezu identische Literaturverzeichnisse aufgestellt hatten, teilweise fanden sich auch in den Quellen keine der erwarteten Angaben. Einige Literaturverzeichnisse waren – vorsichtig gesagt – nachlässig erstellt. Seit dieser Zeit habe ich es als Verpflichtung empfunden, nur das zu zitieren, was ich auch tatsächlich im Original gelesen habe.
Die Aufgabe, die mir Sprankel gestellt hatte, das Harnmarkieren der Riesengalagos zu untersuchen, konnte ich nicht lösen, da das von Eibl-Eibesfeldt [46] beschriebene Verhalten kein Markieren war.
Doch war Sprankel zu einer Themenänderung bereit, Titel der Diplomarbeit war dann
„Die ethologische Bedeutung des Harnwaschens von Galago crassicaudatus E. Geoffroy,
1812 (Lorisiformes: Galagidae)“. Korrekt war dieser Titel – wie wir schon wissen –
nicht, da unsere Galagos tatsächlich der Species Galago garnettii zuzuordnen sind.
Zum Harnwaschen hatte ich geschrieben: „Bei diesem Vorgang neigen sich die
Tiere leicht zur Seite, führen Hand und Fuß an die Harnöffnung (Palma und
Planta9
zum Harnstrahl hin orientiert), fangen den Urin mit der Hand auf und verteilen ihn
durch Aneinanderreiben der homolateralen Hand- und Fußflächen.“ ([234], Seite 2)
Ein anderer Vorgang wird von Eibl–Eibesfeldt beschrieben: „Das Tier hob beim
Urinieren, während es auf drei Beinen ruhte, eine Hinterhand in den Harnstrahl
und ließ die schräg gehaltene Fußsohle berieseln. Dann wischte es eingehend
mit einer und dann mit der anderen Hand über die Fußsohle, so die Hand
benässend“([46], Seite 171). Einen solchen Vorgang konnte ich bei Galago garnettii in
tausenden Beobachtungsstunden niemals beobachten. Leider kann ich in der
Rückschau nicht ausschließen, dass dies darin begründet sein könnte, dass
Eibl-Eibesfeldt eine andere Art beobachtet haben mag. Wahrscheinlicher ist
für mich jedoch, dass Galagos schlichtweg nicht lesen können, könnten sie
es, dann hätten sie das Verhalten – wie von Eibl–Eibesfeldt beschrieben –
gezeigt!
Nach meinen Beobachtungen benötigt ein Riesengalago für seine Bewegungen keine
Harnmarken, die Galagos orientieren sich optisch und bewegen sich mit gezielten
Sprüngen. Die Spuren, die Eibl–Eibesfeldt ([46]) von seinem Riesengalago beschrieben
hat, waren Schmutzspuren, die Riesengalagos hinterlassen, an Stellen, die sie
regelmäßig betreten. Bei dem Harnwaschen fand ich keine Anhaltspunkte für eine
Markierfunktion. Weder das Austauschen des Inventars, der Wegfall eingefahrener
Wechsel noch das Betreten neuer Territorien führten zu einer Zunahme des
Harnwaschens. Die Häufigkeit dieses Verhaltens ist vielmehr abhängig von äußeren
Faktoren. Für genauere Experimente wählte ich zwei Männchen, Moritz und Friedrich,
aus.
Beide Männchen wurden in einem Raum in zwei spiegelbildgleichen Käfigen mit
ständigem Sichtkontakt gehalten (Abb. 3.13). Die Käfige standen auf 0,8 m hohen
Labortischen, um das Beobachten zu erleichtern. Noch unbewußt habe ich hierdurch
die Lebensqualität meiner Versuchstiere erheblich verbessert, da Riesengalgos sehr
ungern den Boden aufsuchen, den Käfigboden haben sie offensichtlich hierdurch nicht
als solchen empfunden.
Moritz10
– im rechten Käfig untergebracht – zeigte während der nächtlichen Aktivitätszeit mehr
Ortsveränderungen als Friedrich. Hierbei entwickelte er verschiedene Varianten
stereotyper Verhaltensweisen. Den drei Varianten war ein Fixpunkt und ein spezieller
Absprungmodus gemeinsam: Moritz hängte sich mit allen vier Extremitäten
mit dem Rücken nach unten an die Decke des Zwischenganges, den Kopf
zur Käfigtür hin orientiert, und sprang dann, sich einen leichten Rechtsdrall
gebend, ab. Dem Sprung konnte ein mehrfaches Umdrehen der Körperachse
vorausgehen.11
Moritz lernte nur langsam, neue Gegebenheiten in den täglichen Aktivitätsverlauf
einzubeziehen. Bisher unbekanntes Futter wurde von ihm kaum angenommen; einige
Fertigkeiten – Öffnen von Eiern, Erdnüssen – wurden von ihm nicht erlernt, andere –
Fischefangen – beherrschte er nur unvollständig. An der Umwelt außerhalb des Käfigs
hatte er wenig Interesse. Zu Aktivitätsbeginn kletterte Moritz auf den Schlafkasten,
wusch mit Harn, putzte sich, rieb die Füße und rekelte sich ausgiebig. Frühestens nach
fünfminütigem Durchführen von Komfortbewegungen zeigte er erste lokomotorische
Aktivitäten. Friedrich dagegen kletterte zu Aktivitätsbeginn zuerst auf den Boden und
begann dort zu fressen und zu trinken. Erst nach der Nahrungsaufnahme
führte er Komfortbewegungen und Rekelsyndrom durch, entweder auf dem
Boden oder im Zwischengang. Unbekanntes wurde von ihm auch manuell
untersucht. Erreichbare Gegenstände wurden an den Käfig heran- bzw. in
diesen hineingezogen und gründlich mit den Händen und/oder durch Bekauen
untersucht. Stereotype Bewegungsweisen konnten bei ihm nicht beobachtet
werden.
Moritz und Friedrich waren also zwei unterschiedliche
Persönlichkeiten. Kurz gesagt, Moritz war „dumm“ und Friedrich
„schlau“.12
Dieser Umstand war aber segensreich für meine Experimente zum
Harnwaschen13.
Moritz traute sich bei einigen Bodensubstratversuchen nämlich nicht, den Boden
des Käfigs zu betreten, so dass ich unter identischen äußeren Bedingungen
ein Versuchstier hatte, das den Boden nutzte und so unweigerlich mit dem
Substrat in Berührung gelangte, und eines, das ungeplant als Kontrolltier diente.
Temperatur und Luftfeuchtigkeit haben erheblichen Einfluss auf die Harnwaschhäufigkeit,
bei hoher Temperatur und unveränderter Luftfeuchte nimmt die Häufigkeit
des Harnwaschens signifikant zu, bei höhere Luftfeuchtigkeit und konstanter
Raumtemperatur dagegen signifikant ab. Diese Beobachtungen führten zu der
Überlegung, man müsse bei unveränderten klimatischen Bedingungen die Feuchtigkeit
der Hand- und Fußflächen manipulieren. Sprankel und ich diskutierten dieses
Experiment und hatten die Idee, den häufig genutzten Käfigboden zu verändern. Wir
entschieden uns für verschiedene Substrate, nämlich Pflaumenmus, groben Sand,
Kaolin trocken, Kaolin feucht, Baumwolltuch trocken und Baumwolltuch feucht. So
wollten wir einmal Hand- und Fussflächen beschmutzen, zum anderen befeuchten bzw.
austrocknen. Die Ergebnisse waren eindeutig, eine Beschmutzung führte zu einer
gewissen Zunahme des Harnwaschens, das Harnwaschen hat also auch einen
Reinigungseffekt. Die Austrocknung von Hand- und Fußflächen hatte einen massiven
Einfluss, besonders eindrucksvoll war dies bei den Versuchen mit dem trockenen
Baumwolltuch. Dieses Substrat hatte für Moritz keine negative Wertung, Friedrich
suchte den Boden sogar häufiger auf als bei anderen Versuchen. „Augenscheinlich
besaß das weiche Substrat positive Wertigkeit: Mehrfach täglich rannte und
hüpfte (biped) er aufgeregt auf ihm herum, sprang hoch, ließ sich auf den
Rücken fallen, drehte und wälzte sich ausgiebig. Dieses Verhalten konnten
wir bei keiner anderen Bodenbedeckung beobachten.“ (Welker [234], Seite
17)
Die Versuche waren erfolgreich, aber auch eine große „Sauerei“. Besonders schlimm
war hier der Einsatz von Kaolin, wurde doch wirklich alles weiß bestäubt, Raum,
Käfige, Versuchstiere. Einen positiven Einfluss hatte nur das Baumwolltuch, das zu
einer echten Verbesserung des Haltungssystems führte, aber auch zu einer extremen
Verschmutzen der hierzu benutzten Laken. Eine nette Kommilitonin bot mir an, diese
zu waschen. Doch ihre Mutter verweigerte die Waschmaschine und wusch die Laken
mehrfach mit der Hand, bevor sie sauber genug für die Waschmaschine waren. Hiefür
möchte ich mich auch hier bedanken.
Nebenergebnis der Versuche waren auch Verhaltensbeobachtungen an Friedrich.
Friedrich kippte nach Aktivitätsbeginn seinen Plastikfutternapf (Durchmesser: 17 cm,
Höhe: 7,5 cm, Leergewicht: 115 g), in dem ich Milch anbot, um und leckte dann die
ausgetretene Milch von der als Bodenbelag dienenden Acrylglasplatte auf.
Bei den Substratversuchen dagegen trank er erst einmal ausgiebig, bevor er
den Napf umwarf. Dies gelang ihm selbst bei dem feuchten Kaolin, er hob
ihn mit beiden Händen an der Unterkante an und drehte ihn sukzessive
hochkant.14
Darüber hinaus beobachteten wir bei Moritz und Friedrich
regelmäßig15
das Fußreiben. Das jeweilige Tier reibt mit der Sohle oder nur der Ferse eines Fußes
intensiv auf dem Substrat (Schlafkastendeckel, Schlafkasteninnenboden, Zwischengang,
Käfiggitter oder (selten) Boden. Dieses Verhalten steht in keinem Zusammenhang mit
dem Harnwaschen. Ein Zusammenhang mit der generellen lokomotorischen Aktivität
ist aufzeigbar. Fußreiben ist zudem laut, diesem Verhalten könnte Bedeutung in
der Fernkommunikation zukommen, so hatte ich mehrfach den Eindruck,
dass auf das Fußreiben mit Fußreiben im zweiten Haltungsraum geantwortet
wird. Sicherlich steht dieses Verhalten auch im Zusammenhang mit einer von
Montagna und Yun [142] beschriebenen Besonderheit des Riesengalagofußes,
nämlich einem bürstenartigen begrenzten Areal mit kurzen hornhäutigen
Stacheln16.
Diese Autoren vermuten, dass diese Region möglicherweise nützlich wäre bei der
Körperpflege und dass diese einen besseren Griff ermöglichen könnte. Für beide
Annahmen fand ich keinen Beleg, kann aber freilich die zweite nicht völlig
ausschließen.
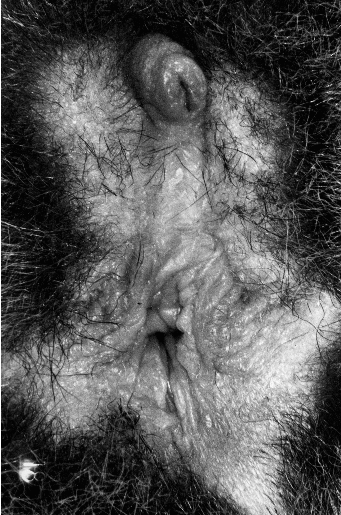
Ich habe schon berichtet, dass ich vier Riesengalagos Anfang Februar 1971 erhalten hatte. Neben Moritz und Friedrich waren dies ein Paar, deren Partner individuell auseinander gehalten werden mussten. Äußerlich war dies einfach, der eine war größer als der andere, doch wollten wir wissen, wer nun das Männchen und wer das Weibchen ist. Sowohl Sprankel als auch ich waren keine Meister im Handeln von Wildtieren. Daher beschlossen wir, nur den größeren der beiden zu „sexen“, ich fing ihn ein und gemeinsam bestimmten wir das Tier eindeutig als Männchen, erkennbar am Penis und am Hodensack. Das zweite kleinere Individuum war demnach das Weibchen. Ich gab den beiden Riesengalagos die Namen Siegfried und Maria. Es sollte dann noch fast vier Monate dauern, bis Siegfried Mutter wurde und ich am 23.05.1971, dem Geburtstag von Tommy, in meinem Beobachtungsbuch notierte: „Siegfried ist Maria und Maria ist Siegfried“.
Auf die Schwierigkeiten, die Geschlechter bei Primaten auseinander zu halten, werde ich noch mehrfach eingehen. Hier schon einmal die Lösung für Maria, ihr Penis war kein Penis sondern die Klitoris, die wie ein Penis aussieht, man kann dies unterscheiden, wenn man darauf achtet, dass der Penis eine Röhre und die Klitoris eine Rinne ist. Wie ein „Hodensack“ sieht die drüsenreiche Umgebung der bis zu eine kleinen Öffnung, direkt unter der Klitoris, zugewachsenen Vaginalöffnung aus. Ich hatte die Eltern vor der Geburt nicht separiert, obwohl dies nach dem Stand der Literatur geboten gewesen wäre, Buettner-Janusch ([13]) betont, wie wichtig die Separation des Vaters sei.17 Entsprechendes berichtete auch Sauer ([181], [182]). Das Absperren schien das Gebot für erfolgreiche Nachzucht zu sein, es war auch die Praxis am Frankfurter MPI. Das Risiko des Verlustes des Neugeborenen ging ich ein, bot aber nach der Beobachtung, dass die Tiere in der Regel getrennt schlafen, drei Schlafkästen an. Der kleine Riesengalago (Tommy) durfte – wie auch weitere Nachzuchten – im Familienverband aufwachsen. Nach der zweiten Geburt (Micky) traute ich mich auch, mit Unterstützung von Dr. Friedrich Jauker die Galagos zu dokumentieren. Micky war zum Zeitpunkt unserer Aufnahmen bereits vier Wochen alt, also relativ selbständig. Dennoch transportierte Maria ihn im Mund bzw. „rettete“ ihn, sobald er Stimmfühlungslaute von sich gab. Nach der Geburt von Hejo hatte ich auch den Mut und das Vertrauen in meine Galagomutter, das heranwachsende Jungtier ab dem ersten Lebenstag für jeweils kurze Zeit zu „entführen“, um es zu wiegen und seinen Entwicklungszustand zu protokollieren und zu dokumentieren. Die wachsende Familie eröffnete so auch Kenntnisse zur Brutbiologie und ermöglichte Befunderhebungen zum Sozialverhalten18.
Riesengalagos werden 50 g schwer
geboren19
und nehmen dann täglich etwa 5 g zu ([232]). Sie sind voll behaart
und haben die Augen geöffnet, diese sind aber wohl noch nicht voll
funktionsfähig20.
Das Junge wird von der Mutter im Mund transportiert. Auf das Mundtragen werde
ich noch in Kapitel 4 ausführlich eingehen. Bereits im Alter von vierzehn
Tagen krabbelt das Junge selbständig im Käfig herum und geht der Mutter
entgegen. Im Alter von etwa vier Wochen nehmen die jungen Riesengalagos
feste Nahrung zu sich. Insgesamt überlebten drei männliche Nachkommen die
Aufzucht im Familienverband, worüber ich auch stolz berichtete ([232]). Richtig
glücklich war ich dann als noch nicht examinierter Nachwuchswissenschaftler,
als die ersten Anforderungen eintrafen, man wollte meine Arbeit tatsächlich
lesen.
Ich hatte nun eine Gruppe Riesengalagos, vier Männchen und ein
Weibchen.21
Die sozialen Interaktionen in dieser Gruppe beruhten auf Kontaktliegen,
sozialer Körperpflege und zwei Formen des Kampfspieles. Das Kontaktliegen
war das Zusammenliegen auf einem der Schlafkästen während der
Aktivitätszeit.22
In der Regel gesellten sich die drei Nachzuchtmännchen Tommy, Micky und Hejo zu ihrer
Mutter Maria, um mit ihr bzw. ihren Geschwistern in engem Körperkontakt zu dösen, dabei
bildeten sie ein enges Knäuel, in dem die einzelnen Tiere nur schwer identifiziert werden
konnten.23
Das gemeinsame Kontaktliegen dauerte meist länger als 15 Minuten an. Für das
Sichputzen nutzen Halbaffen – und so auch die Riesengalagos – neben dem mit einer
Putzkralle ausgestattenen 2. Strahl der Hinterextremität vor allem die Schneide- und
Eckzähne des Unterkiefers, die zu einem Putzkamm ausgebildet sind. Mit diesem
werden die mit den Händen fixierten Stellen des Körpers gekämmt (geschabt). Diesen
Putzkamm setzten sie auch zur sozialen Körperpflege ein. Dabei bevorzugen sie als
aktive Partner die Wangen-, Nacken- und Genitalregion des Partners, als passive
Partner hingegen die Achselregion und die Armaußenseite. Das geputzte Tier streckt
dabei seinen Arm aus und verfällt in eine Putzstarre, die häufig länger als
das Geputztwerden andauert. Das Putzen der Genitalregion führt meist zu
Abwehrbewegungen24.
In dieser Gruppe waren die Männchen die aktiveren und das Weibchen der passivste Sozialpartner bei der sozialen Körperpflege. Sie putzte auch selber seltener als die vier Männchen. An dem Kampfspiel dagegen war sie häufiger beteiligt als alle anderen Gruppenmitzglieder. Das Kampfspiel25 ist die wohl intensivste Form des sozialen Kontaktes, die zwei Varianten lassen sich nach Ort und Art der Entstehung unterscheiden. Bei intensivem wechselseitigen Putzen auf dem Ruhebrett oder einem der Schlafkästen und bei etwa gleicher Putzappetenz wechselte die Rolle des aktiven und passiven Partners in immer schnellerer Reihenfolge. Das jeweils putzende Tier legt sich auf das andere, versucht es festzuhalten und die zu putzende Stelle mit den Händen zu fixieren. Das ständige auf und ab führt schließlich zum Herumkugeln in ventroventraler Position, wobei jedes Tier die gerade erreichbare Körperpartie putzt oder scheinbar beißt.
Sich putzende oder ruhende Artgenossen werden nicht beachtet und oft während des
Herumtollens wie totes Käfiginventar ungezielt berührt. Darauf reagieren diese niemals
aggressiv, sondern versuchen sich zu beteiligen, werden aber jeweils durch Zischen
abgewiesen. Zwischen den zwei Partnern vollzieht sich diese Spielvariante lautlos,
bleibt auf sie beschränkt und kann abrupt von jedem der beiden beendet werden.
Meist putzen sich beide Tiere anschließend ausgiebig. Versuche der Fortsetzung
erlahmen bei mangelnder Antwortbereitschaft eines Tieres schnell. Die zweite Variante
wird nur durch ein Tier hervorgerufen und findet nur auf dem Käfigboden statt. Der
Initiator hüpft biped mit ausgebreiteten Armen herum und wirft sich auf
andere Tiere oder ruft bei ihnen das gleiche Verhalten hervor. Dem dann
gegenseitigen Aufeinanderwerfen folgt wiederum das oben beschriebene Herumkugeln.
Hier können jedoch im Gegensatz zur ersten Variante mehrere Tiere beteiligt
sein.
Zwischen den beiden jüngsten Gruppenmitgliedern, Micky und Hejo, wurden auch
ausgiebige Nachlaufspiele mit wechselnden Rollen – Verfolger und Verfolgter –
beobachtet, auch dieses Spiel konnte von jedem der Partner jederzeit beendet
werden. Nach meinen Beobachtungen waren also die Mitglieder der untersuchten
Riesengalago-Familie ausgesprochen gesellig.
Insofern war ich gespannt, was passieren würde, wenn wir die bisher nur solitär gehaltenen
Männchen Friedrich und Moritz vergesellschaften. Hierüber soll ausführlich berichtet
werden.26
Nach Entfernung des bisher fest installierten Trennschiebers um 20.00 Uhr
blickten beide Männchen sekundenlang – in völliger Körperstarre auf ihren
Schlafkästen verharrend – zum Zwischengang, dann sprang Friedrich ab und landete
in dem Teil des Zwischenganges, das vordem zu Moritz’ Territorium gehört
hatte; Sekunden später folgte Moritz. Beide versuchten nun olfaktorisch die
Genitalregion des anderen zu untersuchen und beschrieben so (immer noch im
Zwischengangsteil von Moritz) beim quadrupeden Hinterherlaufen etwa 30 mal
hintereinander eine enge Kreisbahn. Anschließend versuchte Moritz, Friedrich
zu besteigen und dessen Kopf seitlich zu putzen, während dieser auf Moritz
Genitalregion zielte. Beide wehrten den Putzversuch des anderen ab, indem sie
ihn mit beiden Händen wegboxten und den Kopf mit der Brust zu Boden
pressten. Dadurch bedingt kugelten sie sich minutenlang im Zwischengang herum,
wobei der jeweils obenauf liegende seinen Putzversuch erneuerte. Nach sieben
Minuten sprang Friedrich – gefolgt von Moritz – unvermittelt auf den Kasten
von Moritz, zurück zum Zwischengang und von dort auf den Käfigboden
(Moritz’ Territorium). Dort rannten und sprangen sie schnell und ungezielt
herum, suchten dann wieder den Zwischengang auf und nahmen hier – nun im
gesamten Verbindungsgang – ihren spielerischen Kampf wieder auf. Einander
festhaltend kugelten sie lautlos – Mund und Augen geöffnet, Ohren nicht
gefaltet27
– weitere 37 Minuten umher, dabei vornehmlich Nacken-, Hals- und Kopfregion des
anderen putzend. Da sie hierzu jede gerade erreichbare Körperstelle mit beiden
Händen fixierten, rissen sich beide Tiere in den ersten zwei Tagen nach der Fusion
beträchtliche Haarmengen aus. Bei nachlassender Intensität löste Moritz durch
neuerliches Aufsteigen immer wieder eine Weiterführung des Kampfspieles aus. Um
20.46 Uhr wechselten sie auf den rechten Käfigboden (Moritz) über, hier ihre Balgerei
weitere 55 Minuten fortsetzend. Bedingt wohl durch den – im Vergleich zum
Zwischengang – weiten Raum, traten neue Komponenten auf: Sie hüpften auf- und
übereinander, traten sich mit den Füßen – auch Sohle gegen Sohle gelegt –, warfen
sich auf den Rücken – alle vier Extremitäten nach oben gestreckt –, so den
anderen zum Putzen auffordernd. Nach 22.00 Uhr legten sie kurze Pausen
ein und bewegten sich – einander folgend – in weiten Sprüngen durch beide
Käfige, nun auch den Käfigboden von Friedrich nutzend. Um 22.35 Uhr nahm
Moritz das erste Mal Nahrung zu sich, ohne hierbei von Friedrich gestört zu
werden.
Während der protokollierten vier Stunden des ersten Versuchstages waren beide
Männchen in permanentem Körperkontakt. Die Art der Kontaktaufnahme
erweckt zwar den Eindruck eines Kampfes, entsprach aber dem bereits für die
Familiengruppe beschriebenen Kampfspiel. Der spielerische und friedliche
Charakter der Balgerei war offensichtlich: Es traten keine Verletzungen auf;
gerade das schwächere Tier (Moritz) war meist Initiator; Augen und Ohren
blieben ungeschützt; Fluchtintensionen konnten nicht beobachtet werden. Am
zweiten Tag nach der Fusion zeigten die Tiere ihr Kampfspiel innerhalb der 240
Beobachtungsminuten 177 Minuten lang, eine bzw. zwei Wochen später nur noch 40 -
50 Minuten pro Beobachtungstag. Das Kampfspiel verlief immer lautlos. An den
ersten beiden Tagen nach der Fusion, an denen ich Futter und Milch in beide
Käfige stellte, fand das Spiel vornehmlich im Zwischengang statt. Da die
Männchen keine der Futterstellen bevorzugten, sondern gemeinsam an beiden
fraßen, wurde anschließend nur noch in Moritz’ Käfig Nahrung eingebracht;
dies war wohl Ursache dafür, dass sie nun zum Kampfspiel meist den Boden
von Friedrichs Käfig aufsuchten. In den ersten fünf Stunden nach der Fusion
konnte ein Sichputzen nicht beobachtet werden. Am darauf folgenden Tag
putzten sich Moritz im Mittel 2, Friedrich 1,5 mal in der halben Stunde.
Auch an den weiteren Beobachtungstagen putzten sie sich nur 6,5 mal in 30
Minuten und erreichten somit nicht die bei solitärer Haltung protokollierte
Häufigkeit (Moritz 11,3 ± 0,9, Friedrich 10,7 ± 0,6). Beide Tiere ruhten vor der
Fusionierung bevorzugt auf ihren Schlafkästen (unter einer Rotlichtlampe). Bereits
am zweiten Tag dösten sie nun gemeinsam auf einem Schlafkasten in engem
Körperkontakt. In den Schlafkästen wurden sie jedoch bei täglichen Kontrollen nur
einmal gemeinsam angetroffen (nach einer Woche); eine Bevorzugung des
angestammten Kastens konnte nicht festgestellt werden, schon am Tage nach
der Fusion hatten sie diese getauscht. Vor dem Zusammenlassen waren die
Aktivitätskurven der beiden Männchen verschieden. Nach der Fusion fielen
Phasen hoher und niedriger Aktivität zusammen; an den ersten Tagen und in
jeweils den ersten Stunden der folgenden Tage waren ihre Bewegungen sogar
synchronisiert. Moritz Stereotypie trat am zweiten Beobachtungstag vier mal
hintereinander auf, obwohl Friedrich auf dem in den Bewegungsablauf einbezogenen
Kasten ruhte. Nach einer Woche konnte sie 4 mal in 30 Minuten, nach vierzehn
Tagen 0,4 mal in 30 Minuten beobachtet werden. Sie blieb damit für den
Aktivitätsverlauf ohne Bedeutung und kann als eingestellt betrachtet werden. Bei
zweimonatigen regelmäßigen Kontrollen war das stereotype Verhalten nicht mehr
festzustellen. Andere Verhaltenseigentümlichkeiten blieben unverändert. Schon
am zweiten Versuchstag (um 20.46) warf Friedrich seinen Trinknapf wieder
um.
In den fast zwei Jahren in Gießen erhielten die Galagos täglich Milch (für menschliche
Kinder, frühe Ernährungsstufe) und Bananen angeboten, dazu im täglichen Wechsel
Obst und Gemüse und eiweißhaltige Nahrung (Fische (Schwertträger), Katzenfutter,
Ei roh oder gekocht, tote Eintagsküken, Quarkbrei). Ich fütterte vor Beginn der
Aktivitätszeit und im Überfluss, so dass den Tieren Nahrung über die gesamte
Aktivitätszeit zur Verfügung stand. Streit um Nahrung konnte ich niemals beobachten,
doch war in der Familiengruppe auffällig, dass die Nahrung, die ein anderes
Individuum gerade in den Händen hielt, attraktiver war als die gleiche frei zugängliche
Nahrung.
Sie nahmen dem Artgenossen die Nahrung ab (aus dem Mund oder aus den Händen)
oder knabberten oder leckten am selben Stück, wenn dieser nicht losließ.
„Unbekanntes Futter wird stets vor dem Zubiß optisch fixiert, dann vorsichtig
berochen, betastet und beleckt. Hier machen meist Tommy oder Hejo den
Anfang; erst danach wagen die anderen Tiere, die neue Nahrung zu probieren;
nach vorsichtigem Herumkauen nehmen sie diese nicht immer auf. ... Bei
lebender Beute können die Tiere ihren eingefahrenen (oder ererbten) Modus
der Nahrungsaufnahme nicht anwenden, sondern müssen für jedes Beutetier
gesonderte Fangtechniken entwickeln: Frei umherspringende Heimchen werden nach
kurzer optischer Orientierung zielsicher angesprungen, mit den Händen am
Boden festgehalten und durch Zubiß getötet; größere Orthopteren (Stab-
und Wanderheuschrecken) dagegen werden gepackt und mit beiden Händen
zum Mund geführt. Lebende Fische (Zahnkarpfen), die in einem Wassernapf
schwimmend angeboten werden, fangen die Tiere immer mit den Händen; eine
Unterstützung durch den Mund wurde nur bei Erstkonfrontation mit dem
Medium (Wasser) versucht.“([235], Seiten 57 - 58) Die bewährte Gießener
Diät konnte nicht beibehalten werden und fand schon ein Ende nach der
Vergrößerung der Kolonie um zwei Galago garnettii panganiensis am 15. Oktober
1973.28
Bereits am 17. Oktober fiel mir auf, dass es in dem Haltungsraum nach Pissoir roch.
Die beiden neuen Riesengalagos, Werner und Ilse, waren begeisterte Milchtrinker
und hatten die im Überfluss angebotene Milch durch sich durchfließen lassen. Seitdem
bot ich den Riesengalagos wöchentlich nur noch einmal Milch und nur 50 ml pro Tier
an. Auch das Füttern von rohem Ei (wegen der schwierigen Reinigung und
des geringen Anteils direkt aufgenommener Nahrung) und das Anbieten von
Lebendfutter musste ich aufgeben, da in Kassel nicht an Fischen geforscht wurde,
der stetige kostenlose Nachschub also entfiel. Zudem verfütterte ich keine
Cytrusfrüchte mehr an Galagos, nachdem kurz nach dem Teilumzug der Kolonie
die drei kräftigsten Männchen, Friedrich in Gießen, Siegfried und Tommy
in Kassel an Leberentzündung verstarben. Ein Kollege machte mich darauf
aufmerksam, dass zu diesem Zeitpunkt bestimmte Insektizide verboten und daher
schnell noch im Übermaß verbraucht (also versprüht) wurden. Nach seiner
Meinung könnten spanische Orangen ursächlich für den Tod meiner kräftigen
Riesengalago - Männchen gewesen sein. In der Zukunft und in den nächsten vier
Jahrzehnten erhielten die Riesengalagos weiterhin täglich Banane, dazu im
Wechsel Obst und Gemüse (Apfel, Möhre, Paprika), Katzenfutter, Milch,
gekochtes Ei, Babybrei mit Quark und Lebertran. Der Eiweißanteil war also
hoch.29
Mit zwei mehr oder weniger anekdotischen Bemerkungen soll das Kapitel Gießen
abgeschlossen sein. Zu Moritz und Friedrich nahm ich auch einmal den kleinen Hund
eines Kollegen mit in den Haltungsraum. Sofort näherte sich Friedrich vorsichtig, um
dann wieder zurückzuspringen, diese Form der Annäherung und Erkundung
wiederholte er mehrfach und kontinuierlich. Die Reaktion des Hundes war eindeutig,
näherte sich Friedrich, wich der Hund zurück, sprang Friedrich weg, drängte sich der
Hund bellend vor und versuchte zu jagen. Dem ständigen Wechsel zwischen
Jagen und Fliehen, war er aber nicht gewachsen, er wollte aus dem Raum
raus.
Da ich in der menschlichen Inaktivitätszeit protokollierte, kam es am Morgen zur
Konfrontation mit allen möglichen Geräuschen meiner Mitmenschen, dazu
gehörten auch laute Staubsaugergeräusche und laute Gespräche. Diese störten
meine Galagos nicht, sie dösten z. B. weiter. Im Tierhaus gab es aber auch
rücksichtsvolle Kolleginnen und Kollegen, die das Beobachten respektierten und nicht
stören wollten. Gleichzeitig hatten sie aber auch ein Anliegen. Sie näherten
sich daher extrem leise dem Haltungsraum, klopften leise und flüsterten ihr
Anliegen. Meine Galagos waren sofort hellwach, die Beobachtungssitzung
gestört. Geräusche, die uns „hellhörig“ werden lassen, machen auch Galagos
hellhörig.


1Nach Olson [150] hat Geoffroy seinen Fund - er erhielt das Tier ohne Kenntnis des Fundortes – bereits 1811 beschrieben, diese Veröffentlichung aber nie selber zitiert, wohl weil er ihn als einen Vertreter der Pottos bezeichnet hatte.
2„All the large galagos may now be treated as comprising one widely distributed species under the above name.“ (Hill [85], Seite 217).
3„They are, of course, only local representatives of one single species.“ (Schwarz [198]), Seite 43
4Heute bin ich nicht mehr sicher, ob diese Tiere tatsächlich Galago crassicaudatus argentatus gewesen sind, möglicherweise waren es auch Hybride/Mehrfachhybride von Galago crassicaudatus argentatus und Galago crassicaudatus monteiri, die von Galago crassicaudatus argentatus die Größe des Körpers und von Galago crassicaudatus monteiri die großen Ohren geerbt haben.
6Olson schlägt in seiner Arbeit auch vor, den Riesengalago auf Artenniveau als Otolemur Coquerel, 1859 von den übrigen Galagos zu trennen. Diesem Vorschlag kann ich nicht folgen, da ich den Namen Otolemur als missverständlich für einen Festlandshalbaffen empfinde. Unabhängig davon halte ich Olson’s Ableitung des „richtigen“ Namens für berechtigt.
7The only possible source for Gerrard’s reference to Port Natal which has come to light during this research is that it may be the result of a not completely unjustified misreading of Hoeven (1844: 42)(Olson [150], Seite 128).
8Es sei nicht verschwiegen, dass bereits frühere Chromosomen - Befunde von Egozcue ([43]) und De Boer ([31] darauf hinweisen, dass sich unter dem Namen Galago crassicaudatus verschiedene Species verbergen.
9Heute würde ich statt von Palma und Planta besser von Hand- und Fussfläche berichten.
10Der Text dieses Abschnittes ist nahezu identisch mit dem entsprechenden meiner Dissertation ([235], Seiten 88 -89.
11Das stereotype Verhalten wurde im Mittel mehr als einmal in der Minute protokolliert.
12Ich kann mir das Entsetzen mancher Leser vorstellen, wie kann man einen Galago als dumm bezeichnen, korrekt könnte man auch schreiben, Moritz hatte eine geringere Lerndisposition als Friedrich, doch wird dies Friedrich nicht gerecht. Bei den Galagos ist es eben wie bei den Menschen, es gibt sehr unterschiedliche Intelligenzen.
14Friedrich nutzte den Napf als Spielzeug, das er umherrollte und schob. er transportierte ihn auch mit einer Hand haltend über Kletterstangen auf seinen Schlafkasten. Nach den beschriebenen Versuchsvorhaben, gelang es ihm sogar den Schlafkasten in den Zwischengang zu ziehen, was nur rückwärts kletternd möglich war. Regelmäßig konnte ich dann beobachten, wie er den Napf so lange vorsichtig anstieß bis der Napf mit lautem Krach auf den Boden fiel. Dann holte er ihn wieder und wiederholte seinen Versuch mehrfach.
15im Mittel in dreißig Minuten bei Moritz 8,2 und bei Friedrich 7,4 mal
16„A rough triangular area on the proximal plantar surface, that extends along the tibial margin of the foot to the base of the fifth digit, is covered with short coneal spines that give the surface a brush-like appearance.“[142], Seite 149.
17„The importance of removing the male galago from the cage when the female approaches parturition cannot be overestimated. During the winter and the spring of 1962–1963 exceptional crowding was unfortunately unavoidable in our colony. Three pregnancies terminated in live births. ... In two cases, where the male parent was allowed to remain in the cage, the infants were killed within a short time after birth. The third case, twins born to Hecuba, ..., was more mysterious. Several days after their birth, they vanished without a trace, exactly paralleling the case of the first newborn in the colony.“ ([13], Seite 100)
21Bis dahin wurden nach meinen Recherchen nur Männchen in Deutschland geboren.
22In der Inaktivitätszeit, also am Tage, suchten alle Riesengalagos Schlafkästen auf.
23Scheinbar waren die drei Brüder an der Länge der Schwänze zu unterscheiden, Hejo hatte den längsten und Micky den kürzesten Schwanz. Das Nachmessen ergab, alle drei Schwänze waren gleichlang, der Schwanz von Hejo war schütterer behaart, der Schwarzanteil von Mickys Schwanz war ausgedehnter.
24Abwehrbewegungen sind das „Wegboxen“ mit beiden Händen in das Gesicht des aktiven Partners, Fauchen und Zischen, bzw. die Kombination dieser Verhaltensweisen unter gleichzeitigem Aufrichten des Oberkörpers.
25Die Ausführungen zum Kampfspiel stimmen im Wesentlichen mit denen in meiner Dissertation überein ([235], Seiten 47 - 49).
26Der nachfolgende Text ist nahezu identisch mit demjenigen meiner Dissertation ([235], Seiten 103 - 106).
27Bei einem Kampf sind die Augen und die gut durchblutenden Ohren besonders gefährdet. Daher ziehen kämpfende Riesengalagos ihre Ohren zieharmonikaähnlich zusammen und legen sie an.
28An diesen Transport erinnere ich mich genau, leider nicht an den Namen des hilfreichen Kommilitonen, der mich unterstützte. Ich lieh mir von meiner späteren Frau - wir haben Ende November 1973 geheiratet - ihren fast neuen Sparkäfer und wir fuhren nach München. Auf der Hinfahrt lernten wir, was zuviel Spiel beim Lenken bedeutet und was Aquaplaning ist, der Wagen fährt mit unveränderten Geschwindigkeit und die Tachonadel geht auf Null zurück. In München übernahmen wir die zwei Riesengalagos (Werner und Ilse) und fuhren bei Sturm und starken Regenfällen nach Gießen zurück. Wir - beide junge Männer und begeisterte Autofahrer - kämpften uns zurück und wechselten uns alle dreißig Minuten beim Fahren ab.
29Man mag kritisch einwenden, dass ich nicht versucht habe, die Galagos vegetarisch zu ernähren, doch gab es hierfür keinen vernünftigen Grund.