Abbildung 4.1: Moritz bei dem Aufwachen
1974 war ein überaus erfolgreiches Jahr, sowohl Maria als auch Ilse zogen jeweils ein weibliches Jungtier auf. Ilses Tochter Josephine (30.05.1974) war der erste, Marias Tochter Annemie der zweite (21.11.1974) in Deutschland geborene weibliche Riesengalago. Alle anderen Jungtiere waren Männchen. In Gießen hatte ich gemeinsam mit meinem Doktorvater Heinrich Sprankel darüber spekuliert, welche Faktoren für das Geschlecht der Nachkommen verantwortlich sein könnten und warum gerade nur männliche Riesengalagos geboren wurden. Wir spekulierten über Breitengrade u. ä. Diese Diskussion konnte beendet werden. Nach langjähriger Haltung von Galagos und anderen Primaten kann ich nur berichten, das Geschlechterverhältnis ist immer 1 : 1. Man muss nur eine größere Population betrachten. Dabei treten in der Realität immer wieder auffällige Häufungen von Geburten nur eines Geschlechtes auf, so dass der Halter einmal zu wenige Weibchen, das andere Mal zu wenige Männchen hat. Dies alles ist reiner Zufall. (Mein Freund Gene P. Sackett hat bei einer solchen Diskussion erzählt, dass er einmal bei seiner Zucht von Macaca nemestrina völlig verzweifelt war und händeringend nach plausiblen Gründen suchte, in Folge wurden mehr als vierzig Männchen und kein einziges Weibchen geboren. Auch diese Sorgen waren unbegründet.) Nun waren meine Frau und ich richtig glücklich, ein weibliches Riesengalago - Kind beherbergen zu dürfen.
Josephine wuchs gemeinsam mit ihren Eltern, Werner und Ilse, in einem umgenutzten
Arbeitszimmer in der Hochschule auf, Annemie in unserem Haltungsraum in
Vollmarshausen. Hier konnten wir sie auch für jeweils kurze Zeit zu uns in unsere
Wohnung holen, um sie zu wiegen und ihre rasante Entwicklung vom unbeholfenen
Neugeborenen zum mehrmonatigen Jungtier dokumentieren.
Beide, Annemie und Ilses Tochter Josephine, belegten am Ende des dritten
Lebensmonats die Fähigkeit der Riesengalagos auszubrechen und rechtzeitig
zurückzukehren. Bei Josephine waren es Spuren an der dem Käfig gegenüber stehenden
Schrankwand1,
bei Annemie ein „Verwüsten“ des Käfigvorraumes, die die Vermutung erlaubten,
zumindest einer der gehaltenen Riesengalagos könnte zwischen Außen und Innen
wechseln. Unsere regelmäßigen Kontrollen brachten kein Ergebnis. Immer dann,
wenn wir den jeweiligen Raum aufschlossen, waren alle Tiere im Käfig. In
beiden Fällen brachte ein Trick Abhilfe. Ich schloss die Tür des Raumes zu
und sofort wieder auf und wartete. Das anschließende schnelle Öffnen der
Tür erbrachte die Antwort, Annemie konnte unter der Käfigvorderfront
durchklettern, Josephine durch das Gitter (Maschenweite 5 cm x 5 cm) den Käfig
verlassen2.
Offensichtlich waren beide der Überzeugung, sie müssten im Käfig sein, wenn ich den
Raum betrete, sie hatten eine Vorstellung von „richtig“ und „falsch“. Die Zeit des
Aufschließens (vermutlich auch die Geräusche der Annäherung) reichten aus,
blitzschnell den eigenen Käfig wieder aufzusuchen.

1975 wurde dann als zweites Kind von Werner und Ilse am 24.06. Martin geboren,
ebenfalls ohne Absperren der übrigen Gruppenmitglieder. Was war ich stolz, ich
wusste, wie man Riesengalagos richtig hält.
Wahrscheinlich noch im Jahr 1975 – ich vermute im September oder Oktober – war
ich einmal malad und bat meine Frau, die Galagos zu füttern. Als sie zurückkam,
bejahte sie meine Frage, ob sie denn auch die Türen noch einmal geprüft hätte.
Doch war diese Aussage falsch, am nächsten Morgen war die Tür von Werners
Haltungsraum offen, im Raum befand sich nur Martin. Werner, Ilse und Josephine
waren weg. Josephine fanden wir schnell in dem Installationsraum, in dem
auch das Fenster offen stand. Statt das Fenster zu schließen, packte meine
Frau Josephine mit den Händen und brachte sie trotz heftiger Gegenwehr in
den Haltungsraum zurück. Ilse hatte über das Fenster das Wohnhaus zwar
verlassen, dann aber über ein Kellerfenster wieder aufgesucht. In dem Keller des
Nachbarn war sie leicht einzufangen. Werner fanden wir auf einem Obstbaum
in der Nachbarschaft. Ausgerüstet mit einem Netz und einem Schlafkasten
versuchte ich, durch Schütteln Werner zum Absteigen zu veranlassen. Doch
Werner sprang stattdessen mit einem weiten Sprung auf ein Garagendach und
verschwand dann in den angrenzenden Gärten. Unsere Suche blieb ohne Erfolg.
Wir setzten daher eine Belohnung aus, ich glaube damals DM 500,00, und
veröffentlichten dies in der lokalen Zeitung. Der Erfolg war riesig, die ganze Jugend
Vollmarshausens suchte nach unserem Riesengalago, ständig kamen zwar
falsche Erfolgsmeldungen und Sichtungen, dennoch war die Suche der Kinder
letztendlich ein Erfolg. Am Abend kam ein Junge, der die ganze Zeit besonders
aktiv gesucht hatte, zu uns, der Riesengalago wäre gefunden, in der Wohnung
eines Nachbarn. Sofort machten wir uns auf den Weg und fanden auch schon
Menschen vor einem offenen Fenster im Hochparterre. In diesem Raum wäre er.
Unter Mühen zog ich die Fensterflügel zu und klingelte. Der Wohnungsinhaber
erwartete mich im Flur und verwehrte mir den Zutritt, das Tier wäre über das
Fenster eingedrungen, ich sollte es auch durch das Fenster holen. Nun muss ich
anmerken, dass ich mein Leben lang die lebende Alternative zu sportlich war. In
ein Hochparterrefenster einzudringen, hätte ich mir nie zugetraut. Aber mit
aktiver Mithilfe meiner Frau und wohl anderer Mitmenschen habe ich es dann
geschafft, von außen in das Schlafzimmer einzusteigen. Unter einem großen über
dem Bett angebrachten Kreuz stand Werner und war schnell im Netz. Die
Wohnung durfte ich dann über den Flur verlassen. Wie sich herausstellte, hatte
der Wohnungsinhaber – glücklicherweise vergeblich – versucht, Werner zu
vertreiben, dieser reagierte mit bipedem Aufrichten mit erhobenen Händen und
Drohgeräuschen artgemäß, so dass jener von seinem Vorhaben Abstand nahm.
Die suchenden Kinder brachten die Einsicht, dass vielleicht dieses Tier das
Objekt der Suche sei. Wir haben dann den Finderlohn geteilt, den Hauptteil
erhielt der Junge, der uns auch informiert hatte, für ein neues Fahrrad, ein Teil
drängte ich der Familie auf, zu der Werner sich geflüchtet hatte, für den Rest
veranstalteten wir in unserer Wohnung ein Kinderfest und kauften dazu –
selber noch nicht Eltern – all das ungesunde Süße ein, was Kinder mögen,
mit dem Erfolg, dass einige Kinder anschließend Magenbeschwerden hatten.
Ich hatte bereits berichtet (Kapitel 3), dass ich einer der beiden Familiengruppen und
den einzeln gehaltenen Männchen in Gießen unregelmäßig Schwertträger Xiphophorus
angeboten hatte. Die Tiere lernten, die Fische aus einem Basin zu fangen. Der geneigte
Leser mag einen Selbstversuch mit einem Eimer Wasser und irgendeinem Gegenstand
machen. Das gezielte Greifen ist schwierig, durch den Brechungsindex des Wassers
greift man zu hoch. Umso schwieriger ist dies mit sich bewegender Beute. Diese
Leistung der Riesengalagos wollte ich dokumentieren ([238]). Dazu bot ich der
einen Familiengruppe wieder Fische an, die ich dem Direktor des Genetischen
Instituts der Universität Gießen Prof. Dr. F. Anders verdankte. Zwei der vier
Familienmitglieder, Maria und Micky, kannten bereits Fische, die beiden anderen,
Hejo und Annemie, hatten noch nie lebende Fische gesehen. Offensichtlich
erinnerten sich Maria und Micky an die 18 Monate zurückliegende Futtererfahrung.
Sie näherten sich dem Basin, kontrollierten optisch die Umgebung und das
Becken und griffen vorsichtig in das Basin und rührten mit einer oder beiden
Händen, wohl bis sie einen Fisch berührten, um dann mit schnellem Griff diesen
zu packen, aus dem Wasser zu ziehen und zu verspeisen. Dabei ließen sie
sich durch Fehlversuche nicht entmutigen, sie versuchten es immer wieder.
Die beiden unerfahrenen Tiere, Hejo und Annemie, näherten sich ebenfalls dem
Becken, betrachteten die Fische und versuchten sie mit Hand und Mund gleichzeitig zu
fangen, wobei ihr Mund feucht wurde und sie zurückschreckten. Danach griffen sie
dann nur noch mit den Händen nach den Fischen, ohne Erfolg. Sie untersuchten das
Becken, versuchten an die Fische von der Seite (durch das Glas) zu gelangen
und betasteten das ihnen neue Medium „Glas“. Dann nahm ihr Interesse ab,
gelegentlich suchten sie noch das Becken auf und beobachteten das Fischefangen
durch Maria und Micky. Sie hatten jedoch nicht genug Zeit, die Methode zu
erlernen, da alle Fische von Maria und Micky gefangen und gegessen wurden.

Zur weiteren experimentellen Prüfung boten wir dann der zweiten Familiengruppe ebenfalls
Fische an, jeweils vier Fische an sechs aufeinanderfolgenden Tagen. Während der ersten beiden
Versuchssitzungen3
blieb Ilse in einem der Schlafkästen, Werner beobachtete das Wasserbecken aus
sicherer Entfernung. Am ersten Versuchstag suchte Josephine das Basin bereits nach
zwei Minuten auf und schaute nach den Fischen. Insgesamt suchte sie das Basin
vierzehnmal auf, einmal leckte sie an der Wasseroberfläche. Am zweiten Versuchstag
war das Basin für Josephine nur in der ersten halben Stunde attraktiv, sie suchte es
zehnmal auf. Martin verließ seinen Schlafkasten um 21.40 Uhr, um 22.10 Uhr und
22.14 Uhr näherte er sich jeweils dem Becken und betrachtete die Fische von der Seite.
Beide in Menschenobhut geborenen Tiere machten keine Anstalten, die Fische zu
fangen. Auch am dritten Versuchstag blieb Ilse die ersten fünfundzwanzig Minuten im
Schlafkasten, Josephine und Martin suchten sofort das Basin auf, Werner
nach fünf Minuten. Insgesamt schaute Martin dreimal, Josephine viermal und
Werner fünfmal nach den Fischen. Um 21.55 Uhr verließ Ilse den Schlafkasten
und suchte dann um 21.56 auch das Becken auf. Sofort begann sie, nach den
Fischen zu greifen. Innerhalb einer Minute packte sie nach ihnen elfmal von
der Seite (durch das Glas), dann einmal von oben, einmal von der Seite und
einmal von oben und hatte den ersten Fisch gefangen. Den Fisch im Mund
sprang sie weg, gefolgt von Josephine und Martin, die versuchten, ihr den Fisch
abzunehmen. Um 21.57 Uhr näherte sich Werner dem Basin, griff zweimal von oben
und fing seinen ersten Fisch und sprang mit ihm davon, ebenfalls gefolgt von
den beiden Jungtieren. Im Gegensatz zu Ilse verlor er aber den Fisch an die
Jungtiere. Um 22.04 Uhr fing Werner den zweiten Fisch (ein Griff von der
Seite, zwei von oben), wobei es ihm diesmal durch Abwenden gelang, das
Stehlen zu vermeiden. Um 22.08 Uhr griff Ilse erfolglos achtmal nach dem letzen
Fisch, den sie dann um 22.12 nach zwei Versuchen erbeutete. Während in
Menschenobhut geborene Jungtiere keine Versuche machten, die Fische zu
fangen, lernten dies die Wildfänge innerhalb weniger Minuten. Diese Fertigkeit
verbesserten sie in den nächsten Sitzungen noch weiter, in der vierten fing Ilse
innerhalb von elf Minuten alle vier Fische, in der sechsten innerhalb von drei
Minuten.4
Um zu prüfen, ob auch Nachzuchttiere nach dem Beobachten des Fischefangens diese
Fertigkeit erlernten, mussten wir also die erfahrenen Jäger absperren. Am 6.
Versuchstag boten wir daher um 21.36 Uhr Josephine allein drei Fische an. In den
nächsten 60 Minuten versuchte Josephine immer wieder, von oben Fische zu fangen.
Sie beherrschte offensichtlich das Fischen, machte aber ständig den Fehler, zu hoch zu
greifen ([238]). Dann gab Josephine auf. Nach 45 minütigem Abwarten, testeten wir
diese Vermutung durch Reduktion der Höhe des Wasserspiegels (23.17 Uhr). Sofort
suchte Josephine das Becken erneut auf und fing und aß innerhalb von drei Minuten
alle drei Fische. Galago garnettii sind also in der Lage, lebende Fische zu fangen.
Offensichtlich haben Wildfänge also kein Problem zu erkennen, dass Fische
Nahrung sind. Es muss zuerst eines der Tiere mit den Fangversuchen beginnen,
sie müssen lernen, von oben in das Becken zu greifen, und sie müssen eine
Fangmethode entwickeln, wobei zwei Methoden erfolgreich waren, nämlich
Rumrühren und greifen (Maria) oder schnelles Greifen, ein Griff gefolgt von
dem anderen (Werner und Ilse). Die Wildfänge verbesserten ihre Fähigkeit
von Versuch zu Versuch. Zudem war auffällig, dass das Männchen bereits
bei dem ersten Versuch von oben griff, nachdem es gesehen hatte, wie das
Weibchen einen Fisch fing. In Menschenobhut geborene Riesengalagos müssen
lernen, dass Fische etwas zum Essen sind, offensichtlich lernen sie durch
Zugucken.5
So machte Josephine z. B. nicht den Fehler, von der Seite zu greifen.

Sicherlich wäre es auch interessant gewesen, weitere Lernversuche mit den Riesengalagos zu machen, doch war dies nicht die Zielsetzung meiner wissenschaftlichen Arbeit. In meiner Vorlesung „Einführung in die Ethologie“ war aber das Lernen der Tiere ein wesentlicher Bestandteil. Dabei berichtete ich auch ausführlich über die Versuche des Münsteraner Zoologen Prof. Dr. Dr. Bernhard Rensch ([171]) zum Diskriminationsvermögen verschiedener Tierarten. Meine Studentinnen und Studenten wollten gerne auch solche Versuchsvorhaben mit den von uns gehaltenen Primaten anstellen. Im Prinzip ging es bei diesen Versuchen nicht – wie bei Rensch – um Versuche zur Lernkapazität verschiedener Arten, vielmehr um Fragen, wie plane ich sorgfältig einen Versuch, was muss ich beachten, wie werte ich die Ergebnisse aus. Grundsätzlich eigneten sich unsere Primaten nicht für solche Versuche, da diese zur Motivationserhöhung eine Phase des Nahrungsentzugs beinhalteten, was man bei geselliger Haltung – wie in meiner Kolonie – nicht machen sollte. Dennoch brachten diese Vorhaben viele Einsichten für die Studenten. Bei den Riesengalagos war Annemie das Individuum, das für das Versuchsvorhaben durch den Studenten Jürgen Göllner ausgewählt wurde. Sie sollte zwischen Kreuz (positiv) und Kreis (negativ) unterscheiden, es sollten u. a. Spontanpräferenzen und eine mögliche Seitenstetigkeit kontrolliert werden. Darüber hinaus war bei Erfolg des Vorhabens zu prüfen, ob das positive oder das negative Merkmal gelernt war. Bei solchen Versuchen ist es eigentlich wichtig, die Versuche zeitlich zu begrenzen, um die Versuchstiere nicht zu ermüden. Die in Vollmarshausen durchgeführte Untersuchung erbrachte, dass Annemie ein nicht zu ermüdendes Versuchstier war, sie lernte das positive und das negative Merkmal gleichzeitig6 und beherrschte die zugegebenermaßen nicht schwierige Aufgabe auch Jahre später perfekt.

Im Jahr 1975 erhielt ich auch die Anfrage des Schweizer Künstlerehepaares Jäggli nach einem männlichen Riesengalago. Margrit und Ruedi Jaeggli hatten in ihrem Haus in der Nähe von Bern neben ihren Kunstwerken auch vielen Tieren ein Heim gegeben. Unter diesen waren auch drei Riesengalagos, nämlich Herr Walter, Frau Tobler und Frau Matys.7 Herr Walter war nun einem ebenfalls im Haus gehaltenen Baumozelot Leopardus wiedi zum Opfer gefallen. Konkret wollten sie Ersatz für das Männchen und boten als Tauschobjekt Frau Matys an. Diese lebte in einer befreundeten Familie. Am 05. Januar 1976 war es soweit, Jägglis brachten Frau Matys nach Vollmarshausen und nahmen Hejo mit in die Schweiz.
Wir verpaarten sie mit Micky, was schwierig war, stellte sich doch Frau Matys, eine
sichtlich alte Galagodame, als ungeselliges Individuum heraus. Ich bot ihr jeden Abend
zusätzlich zum Futter einen Teelöffel Beruhigungsmittel an, und – im Nachhinein wie
ein Wunder – Micky wurde am 21.10.1976 Vater, durch seinen Sohn Titus
und dessen Nachkommen war er bei seinem Tod das nach der Zahl seiner
überlebenden Nachkommen erfolgreichste Männchen der Kolonie. Im September
1976 brachte Ruedi Jäggli dann noch Heja, die mit Frau Tobler gezeugte
Hejo-Tochter nach Vollmarshausen. Wir wollten sie mit Micky verpaaren,
doch hatte dieser kein Interesse mehr an weiblichen Riesengalagos, so dass er
mit dieser kämpfte, wobei Heja erheblich verletzt wurde und auch ein Auge
verlor.8
Heja bewährte sich in den folgenden Jahren als Zuchtweibchen, freilich mit einem
anderen Männchen, sie wurde im April 1980 mit einem Enkelsohn von Micky und Frau
Matys, mit Raymund, verpaart.
1976 konnte ich durch das Entgegenkommen der dortigen Kollegen von der Duke
University aus der ehemaligen Buettner–Janusch–Kolonie weitere Riesengalagos
erwerben, nämlich 3 Paare Galago garnettii und ein Paar Galago crassicaudatus, wobei
diese uns als Galago crassicaudatus crassicaudatus bzw. Galago crassicaudatus
argentatus verkauft wurden. Von den sechs Galago garnettii sind zwei Damen
erwähnenswert, nämlich Yoko und Kappa. Die 1972 geborene Yoko hatte schon in den
USA Zwillinge geboren. In Kassel gebar Yoko viermal, davon dreimal Zwillinge mit
unterschiedlichen Vätern. Yoko war also eine echte Zwillingsmutter, was selten bei
Galago garnettii vorkommt. Insgesamt wurden in meiner Kolonie nämlich nur drei
weitere Male Zwillinge von unterschiedlichen Weibchen geboren. Diesen insgesamt
sieben Zwillingsgeburten stehen 97 Einlingsgeburten gegenüber, Galago garnettii
bekommt also in der Regel Einlinge. Kappa ist aus anderen Gründen hervorzuheben,
sie lebte nämlich bei ihrer Ankunft in Kassel bereits mehr als ein Jahrzehnt
in Menschenobhut. Kappa wurde bereits am 19. Oktober 1965 vom Delta
Regional Primate Research Center in Covington von einem Tierhändler erworben
und kam am 07. Januar 1967 in die Primatenhaltung der Duke University.
Als Höchstalter für Riesengalagos galt damals 12 Jahre, dieses Alter hatte
Kappa schon übertroffen, sie war möglicherweise damals der älteste Galago in
Menschenobhut. Als Kappa am 12. Oktober 1980 starb war sie wohl 17 Jahre alt
([244]). Heute weiß ich, dass dieses hohe Alter keineswegs ein Einzelfall ist.
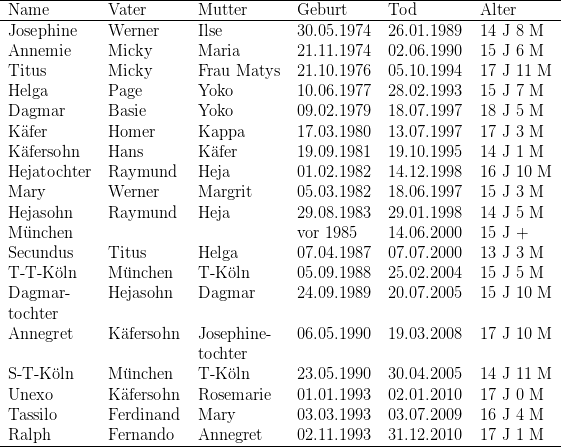
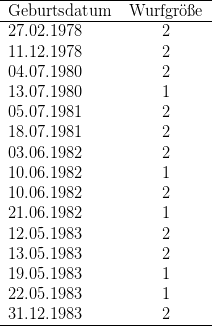
In der Tabelle habe ich alle in meiner Kolonie nachweislich älter als 12 Jahre
alt gewordenen Galago garnettii mit Geburts- und Todesdatum angegeben,
München ist ein männlicher Galago garnettii lasiotis – Wildfang, der nach
Deutschland geschmuggelt, in München beschlagnahmt wurde und am 19.07.1985
nach Kassel kam. Vierzehn Individuen (fünf Männchen, neun Weibchen)
wurden älter als 15 Jahre, acht (vier Männchen und vier Weibchen) älter
als 16 Jahre und sechs (drei Mänchen und drei Weibchen) älter als 17
Jahre. Yokos Tochter Dagmar starb erst im Alter von 18 Jahren und fünf
Monaten.9
Im Sommer 1976 entschied sich auch ein zweijähriger Riesenschnauzer „Poldi vom
Stein“ meine Frau und mich als Frauchen und Herrchen zu adoptieren. Unser
Vermieter in Vollmarshausen forderte mich auf, einen Zwinger zu bauen, doch war
Poldi ein Familienhund und auf den permanenten Kontakt zum Menschen
angewiesen.10
Daher kündigte ich den Mietvertrag zu Ende Oktober 1976 und wir suchten eine
Bleibe, die wir dann in Edermünde–Grifte fanden. In Grifte haben übrigens auch die
erwähnten achtzehn Individuen ihr Höchstalter erreicht. Bedingt durch den
Platzbedarf für unsere Tiere war das Mietobjekt geräumig und reichte auch aus für die
wachsende menschliche Familie. Nach der Geburt unseres Sohnes Armin Ende
November 1976 lernten wir schnell, das Raumangebot in Vollmarshausen wäre nicht
hinreichend gewesen. Poldi haben wir es zu verdanken, dass uns diese Erfahrung
erspart blieb. Bedingt durch die Galagos war der Umzug recht aufwendig, doch halfen
uns Handwerker der Hochschule, besonders der Schreiner Hans Lipp, der die
Haltungsanlage baute, und meine fleißigen Studierenden Barbara Lührmann, Jürgen
Göllner und Marett Grebian, ohne die dieser Umzug kaum möglich gewesen wäre.
Wenn ich 1976 mit der Haltung von Riesengalagos aufgehört hätte, hätte ich nach
fünfjähriger Erfahrung genau gewusst, wie man diese richtig hält. Man biete ihnen genügend
Schlafkästen und genügend Platz und Futter an, dann ziehen sie ihre Jungtiere auch
auf 11.
Ein Abtrennen der Mutter vor der Geburt ist nicht notwendig. 1977 lernte ich, dass die
Aufzucht durch mehrere Mütter bei Galago garnettii nicht funktioniert, dass es also
sicherlich berechtigt ist, die Weibchen vor der Geburt zu separieren. Zumindest Ilse
und Josephine zogen 1977 kein Jungtier auf. Bei beiden gelang eine erfolgreiche
Nachzucht erst 1978 nach Separation. Zahlreiche weitere Beobachtungen führten zu der
Feststellung ([278]), es sei unmöglich, bei mehr als einem Zuchtweibchen von Galago
garnettii ohne Separation Junge groß zu ziehen. Insofern ist erwähnenswert, dass
dies einmal gelang. So wuchs der am 01.01.1993 geborene Unexo, ein Sohn
von Käfersohn und Rosemarie, erfolgreich auf, obwohl Rosemaries Schwester
Annegret, ihr Bruder Fernando und ihre Mutter Josephine-Tochter anwesend
waren.12
Grundsätzlich wurden aber die Mütter von Galago garnettii kurz vor der Geburt
separiert, einige Wochen separat gehalten und dann mit der Restfamilie fusioniert. Die
größte der so entstandenen Familiengruppen umfasste nach meinen Unterlagen neun
Individuen, nämlich Titus und Helga nebst ihren Kindern Primus, Secundus, Tertia,
Quartus, Quintus, Sexta und Septus.
Nach fünfzehnjähriger Haltung war ich sicher, die Mutter-Kind-Beziehung bei
Riesengalagos wird wohl hormonell geregelt. Nach der Geburt sind erstgebärende
Riesengalagos in der Lage, ihre Jungen erfolgreich aufzuziehen, unabhängig von früherer
Erfahrung mit Kindern. Bei Galago garnettii verglichen wir das Pflegeverhalten von sieben
Primipara13,
die nur mit ihrer Mutter bis zur Geschlechtsreife gehalten wurden, dann verpaart und
dann allein gehalten wurden. Sechs der Weibchen (86 % ) zogen den Einling auf,
verglichen mit 8 Multipara, die entweder in Familiengruppen oder mit einem Partner
zusammen lebten und nur vor der Geburt separiert wurden, 12 der 16 Jungtiere (75 %)
überlebten (vgl. [278]). Doch nicht alle Weibchen ziehen ihre Jungen auf, am
03.10.1988 züchtete erstmals die 1982 geborene Mary. Sie pflegte nicht ihr Junges, es
starb nach drei Tagen. Mary züchtete noch acht weitere Male, sie zog keines
der Jungen auf, sechs zogen Studierende und Tierpfleger mit der Flasche
groß.14
Es gibt also auch Riesengalago – Damen, die auch nach Abschluss der Gravidität, also
eindeutig hormonell gestimmt, sich der Aufzuchtarbeit total verweigern. Unter
natürlichen Umständen selektionieren sie sich damit freilich selbst, Mütter mit dieser
Eigenschaft sterben aus.
Die Aufmerksamkeit der Mutter scheint von den Lautäußerungen der
Jungtiere abhängig zu sein. Solange das Jungtier in der arttypischen Weise
mit charakteristischen Klickgeräuschen ruft, antwortet die Mutter mit tiefen
Brummtönen und holt das Jungtier. Gibt das Junge keine Laute von sich,
wird es ignoriert. Auf die Lautäußerungen reagieren alle Riesengalago –
Mütter.15
Die Leistung der Mutter, schnell auf das Junge zu antworten, mag das Töten durch
Artgenossen verhindern. Tatsächlich wurden auch in meiner Kolonie im Laufe der
Jahre einige Kinder getötet und von ihren Verwandten als Beute verspeist. Dies wurde
in allen möglichen Kombinationen sozialer Gruppen beobachtet, bei paarweise
gehaltenen Riesengalagos, Mutter-Sohn und Mutter-Tochter Paaren. Alle
Verwandten, unabhängig von Alter und Geschlecht, waren potentielle Kindertöter.
Zum einen waren meine ursprünglichen Zucht- und Aufzuchterfolge sicherlich
auch dem Umstand zu verdanken, dass meine ersten Galagomütter wehrhafte
Persönlichkeiten waren und meine Galagoväter keine trainierten Killer, zum anderen
wurden die ersten Familien jeweils in einem Raum ohne andere fremde Riesengalagos
gehalten, dies gilt auch für die oben beschriebene Familie von Unexo, dieser wurde in
der Quarantänestation geboren, akustisch getrennt von anderen Riesengalagos.
Grundsätzlich scheinen neugeborene Galagos eher Beute zu sein. Die anderen
Gruppenmitglieder nähern sich vorsichtig und „kosten“, worauf das Jungtier
vokalisiert (klickt), die Mutter herbeispringt und rettet. Hat sie dies mehrfach
gemacht, wissen die Geschwister und der Vater, dass das Junge kein Futter ist. Ist
jedoch ein Gruppenmitglied ein trainierter Killer, tötet es das Junge gezielt,
worauf dieses von allen Gruppenmitgliedern, inklusive der Mutter, gegessen
wird.16
Doch zurück zu den Anfangsjahren der Riesengalagohaltung. Eckhard Prange beobachtete im Rahmen seiner
Staatsexamensarbeit17
in Grifte die schon erwähnte Familiengruppe, bestehend aus Werner, Ilse und ihren
mittlerweile ebenfalls erwachsenen Kindern Josephine und Martin. Es war spannend,
ob diese Beobachtungen meine Gießener Erkenntnisse zum Sozialverhalten des
Riesengalagos (vgl. Kapitel 3) bestätigen würden. Über das Kontaktliegen in dieser
Familiengruppe berichtet Prange: „Beim Kontaktliegen war Werner das attraktivste
Tier der Gruppe, das von allen Mitgliedern der Familie aufgesucht wurde. Auch
Josephine und Ilse waren relativ attraktiv, doch wenn letztere Ausgangspunkt der
Ruhegruppe waren, dann blieb diese auf zwei Tiere beschränkt. In einem solchen Fall
wurde Josephine von Martin und Ilse von Josephine bevorzugt. Martin hingegen wurde
am wenigsten zum gemeinsamen Ruhen aufgesucht. Legte er sich zu einem
bereits ruhenden Tier, so wurde die dadurch entstehende Ruhegruppe für den
Rest der Tiere unattraktiv. Die Reihenfolge, in der sich die Ruhegruppe um
Werner aufbaute, war weitgehend davon bestimmt, welches der Tiere ihn zuerst
aufsuchte. In der Regel war es eines der Weibchen, vor allem Josephine, das sich
zuerst zu ihm legte. Auffällig war, dass Martin fast immer Josephine folgte,
egal, ob diese sich als erstes oder zweites Tier zu Werner legte. Martin ruhte
stets nur kurze Zeit gemeinsam mit den anderen Gruppenmitgliedern und
blieb nur bei Anwesenheit von Josephine länger, Ilse hingegen, die wegen des
häufigen Aufenthaltes im Schlafkasten nur selten am Kontaktliegen beteiligt war,
verweilte aber, wenn sie mit den anderen Tieren ruhte, stets längere Zeit in der
Ruhegruppe.“ (Prange, S. 48) Pranges Beobachtungen zeigten demnach ebenfalls,
welche große Bedeutung das gemeinsame Ruhen im engen Körperkontakt für den
Riesengalago hat. Seine Befunde zur sozialen Körperpflege entsprachen denen, die ich
bereits für Galago garnettii berichtet habe (vgl. Kapitel 3), bedingt durch
die andere Gruppenzusammensetzung und individuelle Unterschiede waren
die beobachteten Sozialpartnerpräferenzen anders. In dieser Gruppe war es
Werner, der häufig allein außerhalb der Schlafkästen während der nächtlichen
Aktivitätszeit ruhte, insofern war er attraktivster passiver Sozialpartner für
seine erwachsenen Kinder, besonders für seine Tochter Josephine. Vergleicht
man die Eltern der Familiengruppe, so ist das Männchen bei der sozialen
Körperpflege aktiver als das Weibchen, das nämliche gilt für Martin und
Josephine, wenngleich auf viel höherem Niveau. Bemerkenswert ist sicherlich
das unterschiedliche Präferenzverhalten der beiden, Martin putzte Josephine
und Ilse mehr als doppelt so häufig wie Werner, Josephine dagegen putzte
Werner bevorzugt. Auch in dieser Gruppe konnte Prange das Kampfspiel (in 36
Stunden 103 Minuten lang zwischen Martin und Josephine und 10 Minuten lang
zwischen Martin und Werner) beobachten. Zum Fußreiben berichtet Prange,
dass es häufig und fast ausschließlich von Werner gezeigt wurde, in seiner
Arbeit vergleicht er den Verlauf der lokomotorischen Aktivität mit der des
Fußreibens, die Kurven sind nahezu identisch. Ich hatte für den Riesengalago eine
dreiphasige Aktivitätsrhythmik beschrieben ([239]), mit dem Hauptmaximum in
der zweiten Hälfte der Aktivitätszeit, Prange bestätigte den dreiphasigen
Verlauf, fand aber das Hauptmaximum zur Aktivitätsmitte. Er vermutet,
dass hierfür der Zeitpunkt der Fütterung (um 24.00 Uhr) verantwortlich sein
könnte18.
Um diese Frage zu klären, ließ ich 1979 Christian Pellmann im Rahmen seiner
Staatsexamensarbeit19
die separat gehaltenen Micky und Heja unter verschiedenen Bedingungen (Futter zu
Beginn bzw. zur Mitte der Aktivitätszeit) über jeweils 36 Stunden beobachten. „Die
Aktogramme der lokomotorischen und nichtlokomotorischen Aktivitäten lassen
bei beiden Tieren eine deutliche Abhängigkeit des Aktivitätsverlaufs vom
Fütterungszeitpunkt erkennen. Bei der Fütterung um 18.00 Uhr zeigen die
lokomotorischen Gesamtaktivitätskurven der Individuen ein hohes Anfangsmaximum,
ein mehr oder weniger ausgeprägtes Mittelmaximum und ein Endmaximum das
zugleich Hauptmaximum ist. Durch die Verlegung der Fütterung auf 24.00 Uhr ändert
sich der grundsätzliche Aktivitätsverlauf nicht, doch zerfällt hier das Endmaximum
in zwei klar erkennbare Teile, eines vor und eines nach 24.00 Uhr. .... Für
die Aktivitätsänderungen bei Verschiebung des Fütterungszeitpunktes sind
drei Komponenten zu erkennen. Das nahrungsappetente Verhalten, also die
Nahrungssuche, ist mit bestimmten Bewegungsweisen verbunden; eine Änderung des
Fütterungszeitpunktes bedingt also eine Änderung der entsprechenden lokomotorischen
Aktivitäten. Das Warten auf Nahrung beeinflusst den emotionalen Status der Tiere,
dies äußert sich in bestimmten Bewegungsweisen. .... Das Rasten der Tiere folgt der
Nahrungsaufnahme; eine Änderung des Fütterungszeitpunktes, also das Vorhandensein
oder Fehlen von Nahrung, bedingt eine Änderung des Ruheverhaltens und somit
eine Änderung der lokomotorischen Aktivität.“ (Pellmann, Seiten 66 - 67).
Pellmann protokollierte auch eine besonders gesteigerte Aktivität zu Zeiten der
Fütterungsvorbereitung, die die Galagos akustisch mitbekamen.
Eckhard Prange war nicht nur ein aufmerksamer Hörer meiner Vorlesung, er arbeitete
auch als Werklehrer im Schuldienst und war insofern der ideale Mitarbeiter für Fragen
der Schlafplatzpräferenz, brachte er doch viele praktische Ratschläge der Umsetzung
ein. Er erfand den mobilen Schlafkasten, bei dem man Vorderwand und Deckel
austauschen konnte, so dass die verschiedensten Varianten zur Verfügung
standen, Schlupfloch, vorne unten, Schlupfloch, vorne oben, Schlupfloch oben.
Zudem konstruierte er Leitern, an denen man den Schlafkasten, bzw. ein
alternativ angebotenes leeres Brett nach oben oder unten wandern lassen
konnte.
„Entscheidenstes Kriterium für die Qualität eines Schlafplatzes ist die Höhe vom
Boden. So werden selbst alte, den Tieren bekannte Schlafkästen gemieden, wenn sie
am Boden stehen. Die Höhe vom Boden ist offensichtlich so entscheidend, dass von
einigen Tieren bereits am ersten Tag der neue Schlafkasten angenommen wird. Tiere,
die sich heimisch fühlen, suchen anfangs jedoch den neuen Kasten nur in
Fluchtsituationen auf. Zum Schlafen wird von diesen stattdessen ein hochgelegener,
ungeschützter Platz gewählt. Für die Höhe vom Boden scheinen die Tiere jedoch keine
absoluten Kriterien zu haben. So ergaben unsere Versuche mit dem solitär gehaltenen
Männchen Micky, dass ein absinkender Kasten an Attraktivität verliert, ein
ansteigender hingegen an Attraktivität gewinnt, unabhängig von der absoluten
Höhe. Hierfür dürfte, neben einer olfaktorischen Komponente, der optische
Eindruck des Tieres, ob ein Kasten sicher oder nicht sicher ist, entscheidend
sein.20...
Neue Schlafkästen sind oft unattraktiv. Hierfür dürfte eine olfaktorische Komponente
verantwortlich sein, da selbst baulich identische neue Schlafkästen gemieden werden.
Besonders auffällig war dies in der Familiengruppe, wo alle neu installierten Kästen
anfänglich nicht zum Schlafen aufgesucht wurden. Alte Schlafkästen hingegen
wurden, in ihre alte Position gebracht, sofort wieder akzeptiert. ... Überlagert
werden alle diese Befunde bei nicht solitär gehaltenen Tieren durch soziale
Beziehungen und Veränderungen in der Sozialstruktur. Unsere in der Familiengruppe
beobachteten sozialen Beziehungen spiegelten sich auch in der Schlafplatzwahl
wider. Werner ist auch als Schlafpartner äußerst attraktiv. Martin sucht,
wie beim Kontaktliegen, vornehmlich Josephine auf.“ (Prange, Seiten 75
-78.) Dies änderte sich freilich als Josephine selber Mutter wurde. Mit ihrem
am 31.05.1978 geborenen Sohn Eckhard wurde sie über nahezu zwei Jahre
(bis zum 25.04.1980) in der Inaktivitätszeit fast immer (97 % der täglichen
Kontrollen) im selben Schlafkasten angetroffen, erst danach schlief Eckhard von ihr
getrennt. Bei diesen Kontrollen war auffällig, dass Martin in der Regel in
demselben Schlafkasten wie Werner angetroffen wurde. Dies mag aber auch
darin begründet sein, dass der Kasten für beide besonders attraktiv war,
es gab jedenfalls keine Hinweise darauf, dass Werner hier gezielt aufgesucht
wurde.21
Die untersuchten Galago garnettii hielten in ihrer nächtlichen Aktivitätszeit
regelmäßig engen Körperkontakt zu Artgenossen, wobei diese aber in meiner
Kolonie stets verwandte oder vertraute Individuen waren, mit diesen vertrugen
sie sich über viele Jahre, teilweise über ein Jahrzehnt. Untersuchungen zum
Sozialverhalten von Galago garnettii anderer Autoren beschränkten sich auf das
Beobachten neu zusammengestellter Gruppen mit einem (Drews 1973 ([38]),
Rosenson 1973 ([176]), Ehrlich 1977 ([45])) oder mehreren Männchen (Tandy
1974 ([211]), Roberts 1971 ([172])). Geradezu kennzeichnend waren das hohe
agonistische Potential von einigen der untersuchten Weibchen und große individuelle
Unterschiede.22
Insgesamt gesehen suggerieren die Untersuchungen den Eindruck,
die nachtaktiven Galago garnettii seien unsozial (Ehrlich [45], Seite
212)23.
Unabhängig davon, dass es richtiger wäre, wenn man statt dem Begriff „unsozial“ besser
„ungesellig“ benutzen würde oder sollte, habe ich – so nehme ich zumindest an –
belegt, dass dieser Eindruck nicht stimmt, meine Befunde – zugegebernermaßen
unter den optimalen Bedingungen in Menschenobhut – erlauben über die
Potenz dieser Art zu spekulieren. Beobachtungen in natürlicher Umgebung sind
schwierig24. Doch
sprechen die Befunde auch hier für eine größere Toleranz zu nachwachsenden Individuen, dies gilt für
beide Geschlechter.25
Es ist sicher erlaubt zu spekulieren, dass diese Beziehungen auch durch Verwandtschaft
bedingt sein könnten.
Männchen präferieren Weibchen als Sozialpartner, diese putzen sie gerne aktiv,
bevorzugt in der Genitalregion, was die Weibchen offensichtlich außerhalb
der Paarungszeit nicht mögen. Sie weisen daher die/das Männchen ab. Die
Eltern in unseren Familiengruppen schliefen in der Inaktivitätszeit in der
Regel in getrennten Schlafkästen, wobei sie höher gelegene bevorzugten. „Die
Sozialstruktur der Riesengalagos beruht auf zwei Komponenten, nämlich einer
Tendenz zur geselligen und einer Tendenz zur solitären Lebensweise. Adulte
Tiere separieren sich aktiv von anderen adulten während eines Teils der
Aktivitätszeit. Jüngere Tiere hingegen bevorzugen den engen Kontakt zu anderen
Individuen während der gesamten Aktivitätszeit. Insofern könnte Galago
garnettii26
ein Modell sein für eine Primatenspecies auf dem „Weg“ vom solitären zum geselligen
Leben. Offensichtlich ist nun aber diese Species diesen „Weg“ bereits weit
vorangeschritten.“ (Welker et al. 1982 [278], Seite 193)
Kopulationen konnten bei Galago garnettii regelmäßig beobachtet
werden.27
Oestrische Weibchen werden von allen geschlechtsreifen Männchen verfolgt; diese
prüfen hierbei ihre Genitalregion geruchlich. Auslöser hierfür könnte der vaginale
Ausfluss während des Oestrus sein. Während des Verfolgens und auch während des
Kopulierens (es finden während eines Oestrus mehrere Kopulationen statt, auch an
einem Tag und mit verschieden Partnern) entsteht kein Streit zwischen den Männchen
(vgl. auch Roberts, 1971 [172]). Häufig werden sie jedoch abgewiesen (mit den Händen
weggestoßen, mit geöffnetem Mund angedroht oder scheinbar gebissen) oder es wird
eine Annäherung durch Pressung der Anogenitalregion gegen den Boden verhindert. In
Kopulationsstimmung hebt das Weibchen jedoch den Schwanz – gerade nach oben
gerichtet – , lässt das Männchen seine Genitalregion untersuchen und belecken und
sich besteigen. Hierzu legt sich das Männchen mit dem Oberkörper auf den Rücken
des Weibchens (das eine Hockstellung einnimmt) und hält sich mit beiden
Händen an den Flanken fest. Dann putzt es die Nackenregion des Weibchens
intensiv oder presst sein Kinn tief in das Fell. Mehrfaches Besteigen geht
der eigentlichen Kopulation voraus, wie von Eaton et al. ([42]) ausführlich
beschrieben.28
Das Weibchen beachtet offensichtlich aufmerksamer die Umgebung als das Männchen,
welches oftmals die Augen geschlossen hält, vgl. auch [1], [42]). Andrew ([1])
beobachtete weiterhin, wie ein Weibchen während des Kopulierens den Mund des
Männchens beleckte. Bemerkt werden soll noch, dass scheue Tiere bei einer Störung
die Kopulation nicht unterbrechen. Kopulationen finden auch am Tage statt.
Auffällig ist auch, dass ein kopulierendes Paar bei dem Kopulieren von anderen
Männchen nicht gestört wird. Da die Kopulationen sehr lange dauern und kaum
abgebrochen werden, konnte ich nahezu alle Kopulationen in der Kolonie auch
protokollieren.
Ich habe bereits berichtet, dass ich 1976 von der Duke University auch ein Paar
Galago crassicaudatus erhalten hatte, nämlich Gram und Eve, die Halbgeschwister
waren. Trotz guter Zuchterfolge konnte ich bei dieser Art Kopulationen bei
regelmäßigen Kontrollen nicht nachweisen, so dass die Annahme erlaubt ist, dass das
Kopulationsverhalten anders ist und dass die kopulierenden Partner sich schnell
trennen können. Die beiden Galago crassicaudatus waren leicht zu unterscheiden,
Gram war silbergrau, Eve schwarz.

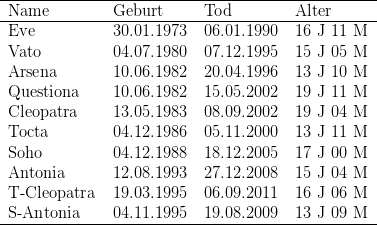
Bei dieser Art sind Schwärzlinge, also die melanistische Form, häufig zu finden. Jungtiere erben entweder die Farbe des Vaters oder die der Mutter. In ein- und demselben Wurf habe ich sowohl graue als auch schwarze Jungtiere gefunden, niemals jedoch intermediär gefärbte. Auch Galago crassicaudatus konnten in meiner Kolonie ein hohes Lebensalter erreichen, das älteste Individuum wurde fast 20 Jahre alt, auch hier habe ich alle Individuen angegeben, die nachweislich älter als 12 Jahre geworden sind, die 1982 geborene Questiona starb kurz vor Vollendung des zwanzigsten Lebensjahres.
Sofort nach dem Kennenlernen von Gram und Eve war mir klar, dass diese sich
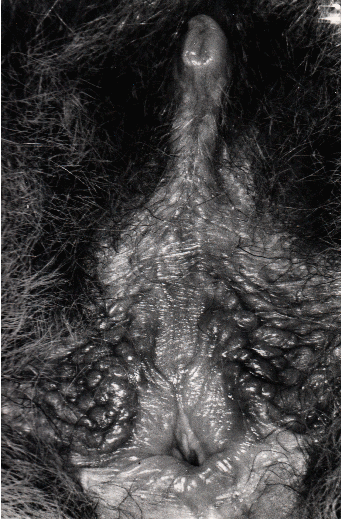
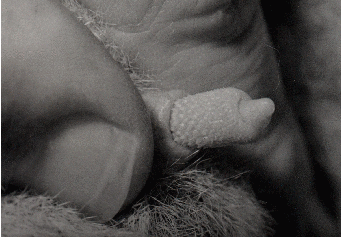
grundlegend von den uns bisher bekannten Riesengalagos unterscheiden. Sie weisen
breitere Ohren auf mit weitbogiger Ohrspitze und sieben Querfalten sowie
einer wulstigen Längsfalte (Plica antitragica) auf. Besonders auffällig sind
bei beiden Geschlechtern die Unterschiede in der Morphologie der äußeren
Genitalien.29
Der Züricher Anthropologe Dr. Wolfgang Scheffrahn untersuchte meine Tiere und
stellte in den Merkmalen des Hautleistensystems eine deutliche Tendenz zur
komplizierten Bemusterung auf, die sich in der erhöhten Bemusterung des kleinen
Daumenballens widerspiegelt. Zudem fand er auffällige serogenetische Unterschiede.
Für mich waren die Verhaltensunterschiede auffällig, im Gegensatz zu allen
Individuen von Galago garnettii, die unabhängig davon, ob sie Wildfänge oder
Nachzuchttiere waren, leicht zu „händeln“ waren, wirkten Gram und Eve
furchtsamer und waren leicht zu stören. Sie bewegten sich schwerfälliger,
zeigten kein elegantes bipedes Hüpfen, auch ihre Territorialrufe klangen
anders.30 Auffällig
war auch ein Drüsenfeld im Hals-Brustbereich, das in dieser Ausprägung bei Galago garnettii nicht
vorhanden war.31
Nach Clark 1982 ([21]) markieren durch Brustreiben dominante Galago crassicaudatus)
häufiger als weniger dominante. An den Marken können sie das Geschlecht des
Markierenden bestimmen, bzw. die Marken verschiedenen Artgenossen zuordnen
(Clark 1982 ([22]).
Zur quantitativen Absicherung dieser Unterschiede im Verhalten ließ ich Gesine
Boehlke32
unter identischen Haltungsbedingungen drei Gruppen Riesengalagos
über die gesamte Aktivitätszeit – insgesamt jede Gruppe 36 Stunden
lang – beobachten, nämlich ein Paar Galago garnettii, ein Paar Galago
crassicaudatus33
und ein Trio Galago crassicaudatus, ein Männchen und zwei Weibchen, Geschwister des Weibchens der
ersten Gruppe.34
Boehlke betont, dass sie das Brustreiben bei Galago garnettii nie beobachtet habe,
das Brustreiben könnte charakteristisch für Galago crassicaudatus sein. „Bei
dieser Verhaltensweise reiben die Tiere ihren Hals-/Brustbereich an einer leicht
rauhen Fläche, sei es eine Leiter, Holzwand oder ein Gitter. Anschließend
erfolgt oft noch ein Reiben des Genitalbereiches und/oder der Hinterbeine in
Fußnähe. Bei dem anschließenden Genitalreiben wird wohl auch ein Urintropfen
abgegeben, somit wäre das Genitalreiben eine Form der Urinmarkierung“ (Seite
135). Zur lokomotorischen Aktivität führt sie aus: „Sicherlich beeinflusst die
Käfigeinrichtung auch die gewählte Fortbewegungsweise. Da die Tiere in nahezu
identischen Käfigen untergebracht sind, können wir diesen Faktor vernachlässigen.
Insofern lassen sich Galago garnettii und Galago crassicaudatus bezüglich
ihrer lokomotorischen Aktivitäten unterscheiden: Galago garnettii ist aktiver,
springt häufiger und zeigt als einzige der beiden untersuchten Species bipedes
Hopsen auf dem Boden“ (Seite 124). Bemerkenwert ist sicherlich auch, dass
Boehlke das Kampfspiel nur bei Galago garnettii beobachten konnte und dass
die beiden Weibchen des Trios mehr miteinander als mit dem Männchen
interagierten. Letztere Beobachtung könnte – wie von Boehlke vermutet – auf
eine höhere Toleranz von Weibchen untereinander bei Galago crassicaudatus
hinweisen, könnte aber auch darauf beruhen, dass die beiden Weibchen Zwillinge
waren.
In der Mutter-Kind-Beziehung treten die auffälligsten Unterschiede zwischen den
Arten auf. Neugeborene Galago garnettii liegen unter oder neben der Mutter.
Neugeborene Galago crassicaudatus dagegen hängen im Fell der Mutter und klettern
auf ihr herum. Die Kinder beider Arten werden regelmäßig im Mund getragen, junge
Galago crassicaudatus bereits vom vierten Lebenstag an auch auf dem Rücken, am
Bauch oder am Schwanz der Mutter bzw. des Tragtieres. Ab dem Ende der ersten
Lebenswoche wehren sich auch die jungen Galago crassicaudatus dagegen, in dem
Schlafkasten abgelegt zu werden. Wegen der häufigen Mehrlingsgeburten – besonders
auffällig war die Tendenz zu Zwillingsgeburten, in der Regel wurden Zwillinge
(23 mal), seltener Einlinge (11 mal) und Drillinge (3 mal), einmal Vierlinge
geboren35
– schienen die Mütter überfordert zu sein, während sie z. B. das zweite Junge
ablegten, klammerte sich das vorher abgelegte in ihr Fell, so dass Galago
crassicaudatus – Mütter häufig ihre Jungen mit herumschleppen. Vergleicht man
Mutter-Kind-Beziehungen bei Galago garnettii mit denen von Galago crassicaudatus
ist man versucht, zu vermuten, dass letztere ein Übergangsmodell darstellen zwischen
Halbaffen, die ihre Jungen normalerweise im Mund transportieren und solchen, die ihre
Jungen am Bauch oder auf dem Rücken tragen.
Offensichtlich ist Galago crassicaudatus noch geselliger als Galago garnettii, so konnte
ich Gram und Eve gemeinsam mit ihren Kindern und Enkelkindern in einer großen
Familiengruppe halten. Die ersten in Grifte geborenen Zwillinge zog Eve nicht auf,
dann überlebten die am 11. Dezember 1978 geborenen Zwillinge Remus und Roma, in
der Regel schliefen diese gemeinsam mit ihrer Mutter, doch traf ich sie bereits in den
ersten Lebensmonaten sechsmal auch getrennt an, ein Jungtier schlief mit Gram, das
andere mit Eve. In der Regel wurden sie in den ersten 461 Lebenstagen gemeinsam
angetroffen (94 %), in den übrigen Fällen schlief entweder Remus (10 mal) oder Roma
(16 mal) mit der Mutter. Bis zum 16.06.1980 schliefen beide in der Regel
gemeinsam mit Eve (86 %). Danach war Eve nicht mehr mit ihren Kindern beim
Schlaf anzutreffen, was sicherlich auch mit der erneuten Geburt von Zwillingen
(04.07.1980) zusammenhängt. Remus und Roma ruhten gemeinsam bis zum 12.
Juli 1980. Am 13. Juli brachte Roma ein Jungtier zur Welt, das aber am
22.07.1980 starb. Ab dem 10.08.1980 bis Ende 1980 (Einstellen der täglichen
Schlafplatzkontrollen) traf ich dann wieder die Zwillinge (Remus und Roma)
stets gemeinsam an, ihre jüngeren Geschwister gemeinsam mit der Mutter.
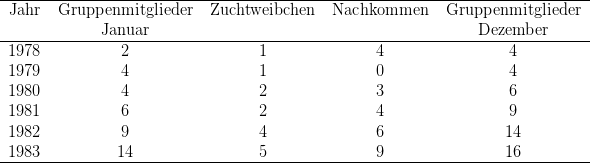
Die Auflistung der Gruppenmitglieder36 belegt die Potenz und die Limitierungen bei dieser Art. 1982 warfen vier Weibchen etwa zur gleichen Zeit. Wenn eines der sechs Jungtiere vokalisierte, antworteten alle Mütter, sprangen herbei und versuchten, das Jungtier zu retten. Daraus resultierten ernste Streitigkeiten zwischen den Weibchen. Gelegentlich konnte ein Weibchen bis zu fünf Jungtiere sammeln, um sie vor den anderen zu schützen. Es wurde auch beobachtet, dass Weibchen Kinder unterschiedlicher Größe säugten, was die Annahme erlaubt, dass Mütter auch zur Aufzucht anderer Weibchen beitragen. Unabhängig von diesen Beobachtungen vermute ich, dass die Weibchen in der Lage sind, zwischen den eigenen und fremden Kindern zu diskriminieren. Am Ende ist es die biologische Mutter, die mit ihren eigenen Kindern angetroffen wird. 1983 zeigten sich - bei weiterer Vergrößerung des Sozialverbandes die Limitierungen, die Zahl der überlebenden Jungtiere ging dramatisch zurück.
Das Verhalten der Weibchen, Junge retten zu müssen, war nicht im Interesse der
Jungen. Weitere Galago crassicaudatus zog ich in diesem Sozialverband nicht auf. Das
Unvermögen der Weibchen, zwischen den Notrufen eigener und fremder Jungtiere zu
unterscheiden, und das Unvermögen der Weibchen, Notrufe ignorieren zu können,
verhinderte größere Geselligkeit. Zudem zeigte sich in diesem Sozialverband, dass es
bei Galago crassicaudatus offensichtlich keinerlei Inzuchtvermeidung gibt. (Nämliches
gilt auch für Galago garnettii.) Dass Galago crassicaudatus noch geselliger ist als
Galago garnettii, wird auch durch Freilanddaten gestützt. Caroline Susan Harcourt
zitiert in ihrer Dissertation (Harcourt 1984 [66]) aus einer eigenen unveröffentlichten
Master - Arbeit und weiteren unveröffentlichen Arbeiten von Bearder, Katsir
und Crompton Berichte zur Geselligkeit von Individuen der Species Galago
crassicaudatus.37
Auch Clark (1985 [23]) gibt Daten einer Feldstudie an Galago c. umbrosus,
nach denen eine hohe Rate direkter freundschaftlicher Kontakte zwischen
erwachsenen Individuen beiderlei Geschlechtes und unterschiedlichsten
Alters beobachtbar sind. Sie berichtet auch Unterschiede zwischen den
Geschlechtern38
und Beobachtungen zur sozialen Körperpflege, die auch den agonistischen Aspekt mit
einschließen39.
Wir werden bei der Besprechung der Affen noch mehrfach auf die soziale Körperpflege
zurückkommen. Man muss sich bewusst sein, dass das Pflegen des Sozialpartners mit
den Fingern bei den Affen eine andere Qualität hat, vergleicht man es mit dem Pflegen
mit dem Zahnkamm bei den Halbaffen. Reinmar Hager beobachtete im Rahmen seiner
Diplomarbeit40
drei Paare von Galago garnettii, wobei zwei Individuen handaufgezogen waren. Er
beobachtete auch das Abwehren der Männchen durch die Weibchen, interpretierte dies
aber als gerichtetes agonistisches Verhalten, worin ich ihm nicht folge. Hager
machte tatsächlich die gleichen Beobachtungen, die ich bereits beschrieben
habe,41
interpretierte diese aber nicht als eigentlich friedliches (ohne Verletzungen) Beenden
der als agonistisch empfundenden massiven Annäherungsversuche der Männchen. Ich
hatte das Verhalten der Weibchen als „Abwehr“ beschrieben und bei der sozialen
Körperpflege zwischen Putzen und Putzen mit Abwehr differenziert. Diese Abwehr
ging „friedlich“ aus.
Dass die jungen Galago crassicaudatus auch am Körper getragen werden,
wurde von Rosenson 1972 veröffentlicht ([175]), tatsächlich aber bereits
1967 von Sauer beobachtet und beschrieben, aber nicht geglaubt und
„wegdiskutiert“.42
Ich nehme an, hätte Sauer seine Beobachtungen an dem Mutter-Kind-Paar über die
relativ kurze Phase von vierzehn Tagen fortgesetzt, wäre ihm aufgefallen, dass bei
Galago crassicaudatus der Transport am Körper üblich ist, so jedoch wirkte sich die
vielfach veröffentlichte Aussage, der Riesengalago trage seine Kinder nur im Mund, wie
eine Zensur aus.
Das Tragen im Mund ist sicher ein ursprüngliches Merkmal der beiden betrachteten
Species. Das Verhalten ist bei allen Galago-Arten und auch bei zahlreichen Arten
madagaskarischer Halbaffen zu finden ([243]). Die Jungen werden zu einem
vermeintlich sicheren Ort transportiert und abgelegt bzw. „geparkt“. Bei den schon
erwähnten Lorisidae hingegen finden wir das Mundtragen nicht, die Jungen werden
stets am Körper getragen, aber bereits – von Art zu Art unterschiedlich – zu einem
frühen Alter „geparkt“. Bei einem Potto Perodicticus potto beobachtete ich dieses
Verhalten bereits am ersten Lebenstag. Die Mutter hing faultierartig an einem Ast und
stupste das Junge immer wieder mit dem Mund und „lenkte“ es so über die Arme auf
den Ast, auf dem das Jungtier wie tot verharrte. Dann suchte sie alleine nach
Nahrung. Das Parken des Jungen ist also - wie das Parken der Galagos - eine
aktive Leistung der Mutter. Durch das Ablegen der Jungen werden diese
vor den theoretischen Gefahren am Körper der Mutter beim Beuteerwerb
geschützt.
Wir werden in den nächsten Kapiteln lernen, dass junge Affen häufig nicht von ihren
Müttern - besonders von erstgebärenden Weibchen - angenommen werden. Unter
natürlichen Bedingungen würden sie verhungern, in Menschenobhut wird in der
Regel der Versuch des Handaufziehens gemacht, der erfahrenen Haltern auch
gelingt. Bei Galago entsteht dieses Haltungsproblem kaum, da - wie schon
ausgeführt - Jungtiere auch gerne aufgegessen werden, so dass es vernachlässigte
Jungtiere nur selten gibt. Unsere „Produziererin“ von Handaufzuchten war
Mary, eine Tochter von Werner und Margrit, geb. 05.03.1982, wobei Margrit,
geb. 12.08.1978, ebenfalls eine Tochter von Werner (und lse) war. Mary zog
keines ihrer Kinder auf, „verschmähte“ diese aber als zusätzliche Nahrung.
Bei ihrem ersten Jungtier, geb. 03.10.1988, hofften wir noch auf einen
Aufzuchtserfolg43,
doch starb es offensichtlich ungenährt nach drei Tagen.
Die beiden nächsten Jungtiere Benjamin, geb. 07.03.1989, und Flummie, geb.
26.08.1989, zogen Barbara Jantschke und Ursula Schäfer, erfolgreich mit der Flasche
auf. Benjamin und Flummie ließen sich problemlos „handeln“, mochten aber
offensichtlich keine anderen Galagos. Sie zogen daher - nach dem Misserfolg des
gemeinsamen Haltens - in getrennten Käfigen nach Grifte um. Dort lebten sie in
unserer geräumigen Diele und durften in den frühen Abendstunden ihren Käfig
verlassen und den Raum erkunden. Es war sicherlich eindrucksvoll, sie bei ihren
Sprüngen bzw. bei dem bipeden Hüpfen zu beobachten, sie hatten aneinander kein
Interesse, kletterten aber gerne auf den menschlichen Sozialpartnern herum, putzten
und markierten diese. Um das Putzen mit dem Zahnkamm auf dem Kopf oder in der
Nähe der Augen zu akzeptieren, muss man schon Galago-Forscher oder enger
Verwandter eines solchen sein. Ich ertrug es mit Sorge, meine Frau mit verläßlicher
Solidarität, meine Kinder „genossen“ die Zuwendung. Erst nach Wochen
schliefen Flummie und Benjamin gemeinsam in einem der Käfige, so dass wir das
Experiment beenden konnten. Richtig „erfolgreich“ war diese Verpaarung zweifellos
nicht, Benjamin und Flummie mochten Menschen, Galagos wurden mehr oder
weniger toleriert. In der Primatenstation starben beide auch relativ jung im
Alter von dreieinhalb Jahren im Rahmen von Neuverpaarungen. Das vierte
Kind von Mary fand ich bei einem Rundgang durch die Primatenstation mit
Studenten am 03.03.1993. Sofort fand sich eine Studentin bereit, den kleinen
Galagojungen großzuziehen. Tassilo wurde uns dann im Alter von drei Monaten
verständlicherweise zurückgegeben. Er solle es gut haben. Tassilo hat mich dann 16
Jahre „begleitet“, andere Galagos mochte er nicht, die ihm angebotenen Galagodamen
verletzte er erheblich, so dass ich mich entschied, ihn nur noch solitär zu
halten.44
Mit unseren Handaufzuchtversuchen waren wir gescheitert, sieht man einmal davon ab,
dass es uns gelungen war, „Menschenfreunde“ heranzuziehen, was aber nicht unsere Absicht
war.45
Daher verfolgte ich bei den übrigen Handaufzuchten eine andere Strategie. Die am
09.01.1995 geborene Mary-Tochter Fussel wurde ebenfalls von Ursula Schäfer gross
gezogen, doch gab ich ihr vom ersten Lebenstag an einen kleinen, noch keine drei
Monate alten mutteraufgezogenen Riesengalago (Albrecht, geb. 26.10.1994)
zusätzlich zur Pflege, so dass Fussel nicht ohne Artgenossen heranwuchs. So
verfuhren wir auch bei dem Mary-Sohn Karl (geb. 05.06.1995). Ursula Schäfer
zog diesen gemeinsam mit Fussel und Albrecht auf. Beide Handaufzuchten
konnten wir später als Galagos halten, insofern war dieser Versuch ein Erfolg.
Einschränkend muss ich aber anführen, dass bei vergleichenden Beobachtungen
und Neuverpaarungen, die Reinmar Hager im Rahmen seiner Diplomarbeit
durchführte46,
Fussel und Karl Verhaltensdefizite bei dem Umgang mit fremden Sozialpartnern
aufwiesen, so dass die Neuverpaarungen abgebrochen werden mussten. Positiv ist zu
berichten, dass Fussel 14,5 Jahre alt wurde und mit Karl am 20.03.1999 bzw. mit
einem weiteren mutteraufgezogenen Männchen am 10.01.2005 erfolgreich züchtete und
ihre Jungen auch selber erfolgreich aufzog. Möglicherweise war der Misserfolg bei der
Neuverpaarung von Fussel auch schwangerschaftsbedingt. Insofern war der
aufwendige Handaufzuchtsversuch von Ursula Schäfer sicherlich ein großer
Erfolg.
Abschließend zu den Berichten über die Riesengalagos möchte ich noch einmal auf die
verschiedenen Unterarten eingehen und Argumente für die korrekte Zuordnung
angeben. Dies erscheint mir wichtig, da auch in dem neuesten Primatenband ([141]
nach meiner Einschätzung Bildunterschriften möglicherweise falsch sind. Ich berichte
also noch einmal kurz über die bei uns gehaltenen Subspecies, nämlich über Galago
garnettii garnettii, Galago garnettii panganiensis, Galago garnettii lasiotis, Galago
crassicaudatus monteiri und Galago crassicaudatus argentatus und hoffe hierdurch
künftigen Galagoforschern das Bestimmen ihrer Tiere zu erleichtern. Schwierig ist
sicherlich, dass bei den Riesengalagos zwischen „großen“ und „kleinen“ Formen
differenziert wird, als kleinen Formen werden dann schnell Garnett‘s Galagos, aus
den großen Galago crassicaudatus, wenngleich der große Galago garnettii
panganiensis47
eindeutig ein Galago garnettii ist. Zudem wird zwischen braunen und grauen
Riesengalagos differenziert, wobei die grauen dann als Galago crassicaudatus
angesprochen werden.

![############################################################################
#Galago##garnettii#lasiotis#ist#-#wie#Galago##garnettii#garnettii#ebenfalls#eine#
#kleinere##ostafrikanische#Form,###die#K#¨orperfarbe#ist#aber##eindeutig#grau.#
#In#Vollmarshausen###erhielt#ich#meine##erste#Galago##garnetti#lasiotis#Dame##
#(Frau##Matys#).###Zudem###¨ubernahm###ich#drei#Tiere##dieser##Subspecies#vom##
#Zoologischen#Garten##K¨oln#(Vater#K#¨oln, Mutter#K#¨oln#und#Onkel##K#¨oln#).#Bei#
############################################################################
#einer#Hybridisierung##mit#Galago#garnettii#garnettii#sahen#alle#Nachkommen####
#wie#Galago###garnettii#garnettii#aus.###Insofern##ist#die#F#¨arbung##von##Ga#-
#lago##garnettii#lasiotis##mit#Sicherheit#nicht#dominant###und##wird#bei##einer#
#Hybridisierung#im##Freiland##verschwinden.##(Freilebende##Tiere achten#bei#ihrer
#Sozialpartnerwahl#nicht#auf#Arten#-#oder#Unterartenzugeh#¨origkeit.##W#¨ahlen#
############################################################################
#sie#einen#Vertreter einer anderen#Art, vermehren#sich#die#Nachkommen###nicht,
#sterben##also#aus.#W#¨ahlen#sie hingegen#einen#Partner#einer#anderen#Unterart,#
#beteiligen#sie#sich#an#der#Evolutionsgeschichte#und#verwirren#m#¨oglicherweise#
#menschliche#Beobachter,##die##ihre“#Tiere#nicht eindeutig#zuordnen#k¨onnen.#)
#Das##genaue#Verbreitungsgebi”et##mu¨sste#man###im##Freiland#noch##finden.##Bei#
############################################################################
#Olson#([150])findet#man##Hinweise##zur#Lokalisation),#die#Ausgangspunkt###f¨ur#
#eine#Suche##sein#k#¨onnten.###################################################](welker94x.png)

![############################################################################
#Die#hier besprochenen#Vertreter von#Galago#crassicaudatus#kommen###ebenfalls#
#in##Ostafrika##vor.###Generell##hat##diese#Art##aber##ein##weitaus#gr¨o#ßeres#
#Verbreitungsgebiet#und#ist auch#im##s¨udlichen#Afrika#zu#finden.##############
############################################################################
#Die#grauen##und##schwarzen##Vertreter#von##Galago##crassicaudatus##werden##in#
#der#Literatur in#Anlehnung##an#Buettner-Janusch##([13#])#nahezu##durchg¨angig#
############################################################################
#als#Galago###crassicaudatus##argentatus##beschrieben,##so#auch###in##eigenen#
#Publikationen.#Sauer#gibt f¨ur#seine#Tiere, die#nach#den#Abbildungen#in seinen
#Arbeiten#([182], [181]) unseren#Tieren#sehr#¨ahnlich#sind,#den#Unterartnamen#
#Galago#crassicaudatus#monteiri#an.##Insofern#k#¨onnte#es#sich#bei#den#als#Ga#-
#lago##crassicaudatus#argentatus#bezeichneten#Individuen##m¨oglicherweise#auch#
############################################################################
#um##Hybride#aus#der#Zucht#der#Duke#University#handeln.#Nach##der#Gr¨oße#des#
#K#¨orpers#sind##sie#eindeutig#Galago##crassicaudatus##argentatus, da#sie#gr¨oßer#
#sind#alsdie#zweiteUnterart,#diewir#gehalten#haben.#Doch##ist#diese#Zuordnung##
#m#¨oglicherweise#falsch.######################################################
############################################################################
#Genaues##wissen#wir#nur##von#unserer#Ausgangspopulation###(Gram##und##Eve,#
############################################################################
#dass#sie#in#der#Kolonie#der#Duke##University#geboren#wurden,##dass#sie#einen#
#gemeinsamen###Vater##haben##(Ngini)#und##unterschiedliche##M#¨utter#(Kahawa##
#und##und##Kilo).#Unsere##Tiere##sahen##sehr#einheitlich##aus,#was#aber##auch#
#durch#Inzucht##hinreichend##erkl¨art#werden##kann.##Unsere##Bestimmung###der#
#zweiten#bei uns#gehaltenen#Unterart#(Galago#crassicaudatus#monteiri)#beruht#
############################################################################
#allein#auf#der#Gr#¨oße#der#Tiere.##Da##die#Individuen##eindeutig#kleiner#sind,#
#m#¨usste die#korrekte#Bezeichnung#Galago##crassicaudatus#monteiri#sein.#Eine#
#genaue##Zuordnung###setzt#das##Wissen##¨uber##die##Herkunft##der#untersuchten#
#Tiere#voraus.###Nach##Olson##([150#])##hat#Galago##crassicaudatus#argentatus#
#nur##ein##sehr#begrenztes##Verbreitungsgebiet##(in##der##N¨ahe##des##Viktoria-
############################################################################
#Sees), Galago#crassicaudatus#monteiri#dagegen##ist#im#ostafrikanischen##Raum##
#weit#verbreitet.##Diese##Einschr¨ankungen###muss##man##bei##der#Beurteilung#
#der#Unterartenzugeh##¨origkeit#beachten.####Bei#beiden##Subspecies##kommen###
#relativ#h¨au#fig#melanistische#Tiere#vor,#was#die#Differenzierung##zwischen#den#
#Unterarten#nicht#erleichtert.###############################################
############################################################################
############################################################################
#Vertreter#beider#Subspecies#sind##weitaus#geselliger#als#Garnett’s#Galagos.#
#So#k#¨onnen##in##einer##Gruppe##auch##mehrere##Weibchen###erfolgreich#z¨uchten,#
#wobei#diese#wohl##verwandt##sein##sollten.#Bei##dieser#Species#werden##in#der#
#Regel#Zwillinge#geboren,#die#auch##ab#einem##bestimmten##Alter#am##K#¨orper#
#getragen#werden.##Haltungsm##¨aßig#gedeihen##Individuen##dieser#Art#unter#den#
############################################################################
#f¨ur Garnett#’s Galagos#beschriebenen#Bedingungen##gut.#####################](welker98x.png)
Bevor ich die Halbaffen hier verlasse, möchte ich meinen Befunden
zur Jugendentwicklung der Galagos eine breitere Basis geben. Die
Insektenfresserahnen48
der Primaten beherrschen ein Parken der Jungtiere - wie oben bei Galagidae
und Lorisidae beschrieben - nicht. Nämliches gilt für das Spitzhörnchen
(Tupaia), das früher entweder den Insectivora oder den Primates zugeordnet
wurde.49
Mein Doktorvater Heinrich Sprankel war der Pionier der erfolgreichen
Spitzhörnchenhaltung. Sprankel ([208]) berichtet: „Wir haben für Tupaia des weiteren
keine Beobachtung über eine Mutter-Kind-Beziehung, die an äffisches Verhalten
erinnerte. Es fehlt ferner jeglicher normale Transport der Jungen durch die
Mutter, auch nach Verlassen des Nestes. (Das Heraustragen aus dem Nest
von lebenden Neonaten, die nicht gesäugt wurden, vollzieht sich nach Art
des Beutetragens, d. h. für gewöhnlich durch wahlloses Zubeißen am Kopf.
Tatsächlich werden dann auch solche Junge gefressen, und zwar stets außerhalb des
Nestes“ (Sprankel 1965 ([208]), Seite 181). Mit Sprankel „... bestehen somit keine
zwingenden Gründe für die Annahme, dass dieses Verhalten von voräffischen
Formen übernommen wurde. Es dürfte sich vielmehr unabhängig von anderen
Säugergruppen innerhalb der Primatenordnung entwickelt haben“ (Sprankel [208],
Seite 182).
Die Betrachtung der Verhältnisse bei Tupaia kann sicher auch nützlich sein, um die
Labordaten bei Galago mit den Freilanddaten zu integrieren. Das japanische
Forscherehepaar Kawamichi führte in Singapur eine Freilandstudie durch und prägte
als Bezeichnung für die tupaiide Sozialstruktur den Begriff „solitary ranging
pair“ ([109, 110]). Was ist darunter zu verstehen? Gehen wir gedanklich von
streng solitär lebenden Tieren aus, dann haben alle Individuen keine Kontakte
untereinander (außerhalb der Paarungszeit) und begegnen sich nur feindlich, in jedem
Terrritorium findet man jeweils nur ein Individuum der entsprechenden Art. Diese
Situation finden wir weder bei Tupaia noch bei den Halbaffen. Vielmehr sind hier
Männchen aggressiv zu anderen Männchen und Weibchen aggressiv zu anderen
Weibchen. Haben nun Männchen und Weibchen die gleichen Ansprüche an die
Größe ihres Territoriums finden wir in ein- und demselben Gebiet jeweils
ein Männchen und ein Weibchen, die das Territorium gemeinsam nutzen,
das „solitary ranging pair“. Bereits bei Tupaia ([109, 110]) gibt es auch die
Situation, dass ein Männchen Anspruch auf ein größeres Territorium stellt und
durchsetzen kann, es hat dann Zugang zu zwei oder drei Weibchen. Die monogame
Struktur ist also polygyn geworden. Übertragen wir die tupaiide Situation auf die
untersuchten Galagos, dann wird erwartbar, dass die Partner des Paares in
Menschenobhut getrennt schlafen werden. In Abhängigkeit zum Toleranzverhalten
gegenüber dem eigenen Nachwuchs wird man auch Mütter mit ihren Kindern
finden. Die Situation in unserer Haltung, dass diese Beziehungen lebenslang
bewahrt bleiben und gelten, mag auch für das Freiland gelten, doch ist hier
die Überlebenswahrscheinlichkeit des Individuums nur gering, was geselliges
Zusammenleben größerer Verbände unwahrscheinlich macht. Der Predationsdruck
durch Reptilen (Schlangen), Vögel (tag- und nachtaktive Greifvögel) und Säuger (u. a.
Katzen und Schleichkatzen) ist bei so kleinen Säugetieren (wie Tupaia) bekanntlich
groß.
Da die Beschäftigung mit Jungen ablegenden Halbaffen zwangsläufig dazu führt, dass
man diese mit Halbaffen und Affen, die ihre Jungen am Körper tragen, vergleicht
und im Hinblick auf die aktuelle Diskussion über das korrekte Zitieren von
Quellen, möchte ich hier noch auf den Ontogenese-Zustand der Primates
eingehen.50
Hierzu führte Portmann (1962) aus: „Die Jungen der Primaten durchlaufen
ausnahmslos intrauterin das Stadium des primären Nesthockers und schreiten zur
morphologischen Stufe des sekundären Typus fort, der mit offenen Sinnespforten
und weit entwickelter neuromuskulärer Organisation zur Welt kommt und
dem Nestflüchter der Huftiere und Wale entspricht“ ([165] nach [167],
Seite 288). Die Primaten werden von Portmann dem Ontogenese-Typ des
Nestflüchters zugeordnet. Neben Tupaia, dessen Nesthockersituation unbestritten
ist, gesteht Portmann (1959) nur noch dem Menschen eine Sonderstellung
zu. „Unser Entwicklungsgang weicht in wichtigen Zügen von dem höherer
Eutherien51ab.
Der Mensch durchläuft im Mutterleib in der Zeit vom zweiten bis etwa zum fünften
Fötalmonat zunächst das Stadium der primären Nesthocker mit dem Verschluss der
Sinnesorgane. Er tritt darauf in das Nestflüchterstadium mit offenen Sinnesorganen
ein, in dem er auch zur Welt kommt. Doch entspricht sein Geburtszustand nicht dem
der höheren Säuger: im Gegensatz zum neugeborenen Huftier, Seehund, Wal oder
Makaken zeigt der neugeborene Mensch weder die arttypische Körperhaltung noch die
adulte Bewegungsweise. Dieser eigenartige Zustand muss als sekundärer
Nesthocker52
bezeichnet werden und steht völlig allein unter den Säugern“ ([164], Seite
288). Die Beschreibung des Geburtszustandes des Menschen als sekundärer
Nesthocker und die daraus ableitbare Sonderstellung von Homo innerhalb der
Primates kommt dem Bedürfnis von Homo, sich selber eine Sonderstellung
zuzugestehen, sicherlich entgegen und hat daher weithin Anklang gefunden.
Portmann selber hat nach der Geburt des Gorilla-Mädchens Goma 1959 den
Begriff nicht mehr gebraucht, da der Geburtszustand des Menschen nun ganz
anders erscheint (Portmann 1964): „... trotz seiner Hilfslosigkeit kommt das
Menschenkind mit offenen Sinnesorganen und einer weit fortgeschrittenen
neuromuskulären Ausformung zur Welt. Unser Geburtszustand ist weit vom primitiven
Nesthocker entfernt auf dem Weg zum Nestflüchter – auf dieser Wegstrecke wird er
früh geboren“([166] nach [167], Seiten 328 - 329). Darüber hinaus verkennt
Portmann in seinen späteren Arbeiten auch nicht die vermittelnde Stellung der
Menschenaffen, betont aber weiterhin die Sonderstellung des Menschen. „Die
Abweichung unserer Ontogenese von der typischer tierischer Primaten hat zu
einer Gliederung der Entwicklungszeit geführt, in der die fötale Zeit radikal in
zwei Perioden geteilt ist: 1. in eine typische Uteruszeit, 2. in eine ’soziale
Uteruszeit’, in der die Gruppe die Rolle eines neuen Uterus übernimmt. Die
Anthropoiden53
zeigen Anklänge an eine solche Gliederung“ (Portmann 1962 [164] nach [167], Seiten
291 - 292).
Obwohl Portmann selber in seinen Arbeiten den Begriff „sekundärer Nesthocker“ nicht
mehr benutzte, wurden als Quellen für den „sekundären Nesthocker“ in der Literatur
Portmann 1962 und Portmann 1964 angegeben. Als junger Student habe ich daher
stundenlang nach diesem Begriff in den zitierten Büchern gesucht. Eindeutig waren
diese Zitate Fehlzitate.
Ich hatte und habe meine Zweifel, ob Begriffe wie Nesthocker und Nestflüchter – von den
Vögeln entlehnt – überhaupt sinnvoll für die Bezeichnung des Ontogenesezustandes der
Primaten verwendet werden können. In den nächsten Kapiteln werde ich
Befunde von und Überlegungen zu verschiedenen Arten der Echten Affen
vorstellen.
1Der Büroraum wurde später von Prof. Dr. Helmut Hofer als Arbeitszimmer genutzt. Immer, wenn ich ihn besuchte, ging mein Blick an die Schrankwand, und ich kontrollierte, ob Josephines Spuren noch zu sehen waren, Josephine mochte es, an der oberen Kante des Schrankes entlang zu laufen. Diese Spuren blieben erhalten. Ich habe freilich auch niemanden auf die Spuren aufmerksam gemacht.
3Die Sitzungen begannnen jeweils um 21.30 Uhr, eine Versuchssitzung dauerte maximal 60 Minuten, sie endete vorzeitig, wenn alle Fische gegessen waren.
4In der fünften Sitzung trennten wir Ilse ab, Werner fing alle Fische innerhalb von 15 Minuten.
5Diese Annahme wird durch umfangreiche Experimente von Watson und Mitarbeitern, die mit 38 Individuen meine Untersuchungen wiederholten ([230]), gestützt.
6Man testet dies mit einem oder mehreren zusätzlichen Merkmalen, hat das Versuchstier das positive Merkmal gelernt, wählt es dieses vor einem fremden, hat es das negative Merkmal gelernt, wird bei der Wahl zwischen einem neuen und dem negativen Merkmal das neue gewählt.
7Die Tiere wurden wegen bestimmten Ähnlichkeiten nach lebenden Personen benannt.
8Heja musste tierärztlich behandelt werden, hier sei unserer Tierärztin Dr. Doris Becker herzlich gedankt, und bekam auch zum Schutz einen Kragen, der das Putzen mit dem Zahnkamm verhinderte, gleichzeitig verhinderte der Kragen das Beißen zur Abwehr. So übernahm meine Frau das Putzen von Heja. Sie rechnete nach dem Entfernen des Kragens mit Abwehr, doch ließ sich Heja weiter gerne bei der Körperpflege unterstützen, Heja und meine Frau putzten gemeinsam, Heja mit dem Zahnkamm, meine Frau mit Kamm und (Zahn-)Bürste.
9Über die Namen könnte man streiten, doch habe ich der Versuchung widerstanden, diese im Nachhinein umzubenennen. Aus Köln erhielt ich drei Galago garnettii lasiotis, die wir Vater Köln, Mutter Köln und Onkel Köln genannt haben. T-Köln ist die Tochter von Vater und Mutter Köln, ihre Tochter erhielt den Namen T-T-Köln, ihr Sohn den Namen S-T-Köln.
10Poldi war aggressiv zu Rüden, die ihn dominieren wollten, friedlich aber ansonsten zu Mensch und Tier. Kaum lebte Poldi bei uns in Vollmarshausen, entkam Moritz - und wir fürchteten um sein Leben. Schnell sperrten wir Poldi in das Arbeitszimmer meiner Frau und suchten sorgfältig die ganze Wohnung ab, ergebnislos. Als endlich einer von uns auf die Idee kam, Moritz könnte im Arbeitszimmer sein, schauten wir nach. Poldi lag friedlich unter dem Schreibtisch und Moritz saß auf diesem.
12Einschränkend muss auch angemerkt werden, dass Unexos Großmutter Josephine-Tochter selber am 15.12.1992 Zwillinge gebar, aber nicht aufzog. Sie war wohl selber hormonell auf Pflegen eingestellt, dies mag das Überleben von Unexo begünstigt haben, vgl. auch [232].
13Erstgebärende
14Auf die Problematik von Handaufzuchten werde ich im nächsten Kapitel ausführlich eingehen und hier auch über die Erfahrungen mit Galago berichten. Hier sei nur erwähnt, dass das Handaufziehen ein Fehler ist.
15Unabhängig von der Artzugehörigkeit.
16Neugeborene Riesengalagos scheinen besonders gefährdet zu sein. Nach Experimenten von Rosenson ([174]) sind ältere Jungtiere weniger gefährdet bei Konfrontation mit einem nichtverwandten erwachsenen Artgenossen.
17Prange, E.: Zeitliche Ordnung des Verhaltens und Sozialverhalten in einer Familiengruppe des Riesengalagos Galago crassicaudatus E. Geoffroy, 1812, nebst. Experimenten zur Qualität des Schlafplatzes. Wissenschaftliche Hausarbeit zur Ersten Staatsprüfung für das Lehramt für die Mittelstufe. Kassel, 15. September 1978.
18In Gießen hatte ich die Riesengalagos stets vor Aktivitätsbeginn gefüttert und ihnen auch Nahrung im Überfluss angeboten. Nach jahrelanger Haltungserfahrung wusste ich, welche Futtermengen ein Riesengalago benötigt und bot nur diese Menge an. Da die individuelle Futteraufnahme sehr unterschiedlich war, manchmal nahmen die Tiere tagelang kaum Nahrung zu sich, manchmal vertilgten sie das gesamte angebotene Futter in wenigen Minuten, bot ich ihnen die neue Nahrung zu Beginn der zweiten Hälfte der Aktivitätszeit an. Bis zu diesem Zeitpunkt stand ihnen auch der Futternapf des Vortages zur Verfügung.
19Pellmann, C: Der Fütterungszeitpunkt als „Zeitgeber“ bei dem Riesengalago Galago crassicaudatus. Wissenschaftliche Hausarbeit zur Ersten Staatsprüfung für das Lehramt für die Mittelstufe, Kassel, 10. April 1979.
20Zu Beginn der Versuche boten wir Micky ein Brett 160 cm über dem Boden und einen Kasten mit Schlupfloch (vorn unten) 25 cm über dem Boden an. Dieser Kasten blieb bis zu einer Höhe von 100 cm unattraktiv, wurde dann aber bei derselben Höhe angenommen (besonders nach einer gewissen Adaptationszeit), als der Zwischenraum zwischen Winkel und Kasten mit einer Abdeckung verkleidet war. Kasten und Winkel boten somit eine optische Einheit. Bei einer Absenkung auf 85 cm verlor der Kasten an Attraktivität und wurde in einer Höhe von 70 cm schließlich gemieden. Das anschließende Ansteigen des Kastens auf 85 cm erbrachte jedoch ein ganz anderes Ergebnis. Hier erreichte der Kasten eine sehr hohe Attraktivität, die er auch dann nicht verlor, als die Abdeckung des Zwischenraumes entfernt wurde. Bei einer eneuten Absenkung auf 70 cm verlor der Kasten erwartungsgemäß wieder seine Bedeutung als Schlafplatz und blieb beim Ansteigen auf 85 cm und sogar 100 cm unattraktiv. Dies änderte sich erst, als wir die Zwischenraumabdeckung erneut anbrachten.
21Unfreiwilliges Nebenergebnis der langfristigen Kastenkontrollen war das Faktum, dass ich nach tagelangen Kontrollen und Vorfinden der Individuen in ein- und demselben Schlafkasten sicher war, sie schliefen „immer“ in diesem Schlafkasten. Durch das tägliche Protokollieren waren solche „Zeugenaussagen“ schnell falsifiziert.
22A primary factor in galago social behaviour may be individual preferences and aversions among the animals. ( [172], Seite 181
23These results are in keeping with the generally-held view that nocturnal prosimians are relatively unsociable ...
24Though the G. garnettii individuals generally foraged alone, as do all galagos the radio tracked individuals were occasionally seen with other animals during the night. However, it was extremely difficult to see the bushbabies. Even when it was known that more than one animal was present, identification of any but the collared animal was usually impossible. (Harcourt ([66]), Seite 123)
25Adult females of different ages shared highly overlapping ranges, while like-aged females showed little range overlap. Females matured and had their first infants in their natal ranges. Adult males’ ranges were larger than those of females and overlapped them extensively. Resident adult males showed little range overlap with each other, unless they were of different ages. (Nash und Harcourt ([145]), Seite 357)
26In der zitierten Publikation ([278]) ist der Artname Galago agisymbanus angegeben.
27Die Ausführungen zum Kopulationsverhalten stimmen im wesentlichen mit denen in meiner Dissertation überein ([235], Seiten 16 - 17).
28„Once the male had succeeded in obtaining a clasp mount, he usually remained motionless for a few seconds and then either dismounted or started to thrust. After initiating pelvic thrusts and achieving an erection, he continued thrusting for 30 to 80 s and then dismounted if penetration did not occur. This sequence was repeated until intromission was achieved. After intromission, the rapid shallow pelvic thrusts were replaced by slow (two to ten per min) but vigorous thrusts lasting 4 to 8 min. Vaginal smears showed that ejaculation occurred immediately after the cessation of these deep, convulsive thrusts, not before. The animals then remained quietly, the male resting his chin on the female’s back and maintaining an intromission for up to several hours.“ ([42], Seite 311)
29Diese Unterschiede werden bereits 1977 von Dixson und Van Horn berichtet, sie diskutieren auch die Notwendigkeit der Zuordnung zu verschiedenen Species ([36]).
30Clark 1988 [24] untersuchte die „long calls“ von G. garnettii und Galago crassicaudatus und belegte die Unterschiede, die Tiere antworten häufiger auf Rufe von Argtenossen als auf Rufe von Individuen der anderen Art.
31Das Brustmarkieren war bei Galago garnettii nie sehr auffällig, das Männchen markierte wohl sein Weibchen bei der Kopulation, mich selber markierte ein handaufgezogenes Männchen intensiv auf dem Kopf.
32Boehlke, G.: Vergleichende Verhaltensuntersuchungen an Galago crassicaudatus und Galago garnettii. Wissenschaftliche Hausarbeit zur ersten Staatsprüfung für das Lehramt für die Mittelstufe und die Oberstufe. Kassel, 25. September 1987.
33Das Männchen stammte von einem Privathalter und wurde seit Juli 1983 in meiner Kolonie gehalten, das Weibchen wurde im Zoo Berlin geboren und lebte, gemeinsam mit seinen Geschwistern, seit dem 31. Juli 1979 in meiner Kolonie.
34Diese Tiere waren „anders“ als die Galago crassicaudatus, die wir von der Duke University erhalten hatten. Wir hatten sie als Galago crassicaudatus monteiri geführt und bestimmt [282].
35Einschränkend muss ich darauf aufmerksam machen, dass alle Galago crassicaudatus von Gram und Eve abstammen. Nach den Befunden von Eaglen und Simons [41] kann dieser Einwand aber vernachlässigt werden, bei 60 Geburten in neun verschiedenen Matrilinien von Galago crassicaudatus wurden dreißigmal Mehrlinge geboren, in zwölf verschiedenen Matrilinien von Galago garnettii dagegen bei 86 Geburten nur drei Zwillingsgeburten.
36In den betrachteten sechs Jahren habe ich nur ein erwachsenes Individuum durch Unfalltod verloren.
37In this species there are three categories of males: resident, dominant males with territories overlapping those of more than one female (the size of a male’s range has not been measured); resident, peripheral males which are tolerated by the dominant animals; and vagrant males which are young animals possibly looking for a territory. Adult females also have territories (of about 7 ha) which they often share with female relatives but from which they exclude other females. During the day females and their offspring frequently sleep together while the adult males tend to sleep alone. (Seite 112)
38Thus males seem to disperse farther and may move later in their lives than females (Seite 585).
39Grooming was an important aspect of positive interactions. Allogrooming (unidirectional) occurred in 250 (or 42 %) of such interactions, and mutual grooming (bidirectional) in 295 (49 %). Grooming occurred in sexual (12 %), ambivalent (70 %), and even agonistic interactions (2 %). In the latter cases, the grooming (between males in the mating season) was rough and unidirectional although unmistakable by the forward-and-up head motions made while using the tooth comb. The recipient drew back and the groomer effectively forced the other to run or retreat (Seite 588).
40Hager, R.: Behavioural differences between the sexes in captive Garnett’s greater bushbabies (Otolemur garnettii) under different experimental conditions. Diplomarbeit an der Universität Würzburg. Würzburg, 28. Juli 1999
41The fact that agonistic behaviour in established pairs was virtually only initiated by females is a result of the males’ permanent attempts to sniff the genitals. ... The females did not appreciate this and if the male was too obstinate she reacted with agonism. (Hager, Seite 59)
42„The unusual jockey-style ride of the eight-day old infant on the back of his mother (....) occurred only once in an exceptional situation when the female, in a momentary arousal of fear, left the nest too suddenly.“ ([181], Seite 137)
43Wir wollten auch unseren in den nächsten Kapiteln ausführlich beschriebenen Misserfolgen bei echten Affen nicht noch weitere bei Halbaffen hinzufügen.
44Auch die solitäre Haltung war nicht einfach. Die Türen meiner Galagokäfige hatten etwa „normale“ Türgröße, was sich bei der Haltung bewährt hat. Öffnete man diese, so sprang Tassilo sofort heraus auf den Menschen, also in der Regel auf mich, um mich zu pflegen bzw. von mir aus, die Umgebung zu erkunden, was mit erheblichen Gefahren verbunden war. In allen benachbarten Gehegen lebten fremde Galagos, die zumindest theoretisch bereit waren, die Finger von Tassilo zu „kosten“. Insofern musste ich in Tassilos Tür eine kleine Futterklappe einbauen, die das Füttern ohne Entweichen ermöglichte. Unter diesen Haltungsbedingungen wurde Tassilo alt, er „wartete“ - anthropomorph betrachtet, über Jahre auf die unregelmäßigen Besuche meines Sohnes Armin, der mir - war er in Grifte - bei der abendlichen Fütterung assistierte und Tassilo’s Tür öffnete. Tassilo wartete auf diese Chance, er genoss es offensichtlich, in Armins langen Haaren herumzuturnen und ließ sich auch immer problemlos in seinen Käfig zurücksetzen.
45Gelegentlich musste ich auch die Galagos handeln, dabei ist es sehr wichtig, sie richtig festzuhalten. Man spürt dann auch den Herzschlag des festgehaltenen Riesengalagos. Danach ist das Handeln keine große Belastung. Anderes galt für Jungtiere und für Tassilo. Erstmals eingefangene Jungtiere sind sehr aufgeregt, bei Tassilo hingegen konnte man den Herzschlag kaum spüren, er genoss es offensichtlich, festgehalten zu werden.
46Hager, R.: Behavioural differences between the sexes in captive Garnett’s greater bushbabies (Otolemur garnettii) under different experimental conditions. Diplomarbeit, Universität Würzburg, Juli 1999.
47Unsere Tiere wurden nach Dennig ([35]) in Tanzania, in der Nähe von Arusha gefangen.
48Zur Ordnung der Insektenfresser, der Insectivora, gehören u. a. Spitzmaus, Maulwurf und Igel
49Die vielen gemeinsamen Merkmale, die Tupaia mit den Primates teilt, sind ursprüngliche Merkmale und taugen daher nicht zur Konstruktion von Verwandtschaft. Heute werden die Spitzhörnchen einer eigenen Ordnung Scandentia zugeordnet. Die Einsicht, dass es wichtig ist, zwischen ursprünglichen und abgeleiteten Merkmalen zu differenzieren (bzw. zwischen „plesiomorph “und „apomorph“, verdanken wir wiederum Willi Hennig.: „We will call the characters or character conditions from which transformation started (a, b) in a monophyletic group plesiomorphous , and the derived conditions (.......) apomorphous. Simple reflection shows that these are relative concepts ...... “ (Seite 89) „We will call the presence of plesiomorphous characters in different species symplesiomorphy, the presence of apomorphous characters synapomorphy, always with the assumption that the compared characters belong to one and the same transformation series. “ (Seite 89). „It is evident that the presence of corresponding characters in two or more species is a basis for assuming that these species form a monophyletic group only if the characters are apomorphous, if their correspondence rest on synapomorphy.“ (Seite 90)
50Die Fragen und Gedanken zum Ontogenese-Zustand der Primaten haben mich schon in der Gießener Zeit und bei der Arbeit an meiner Dissertation beschäftigt. Sehr viel Mühe hatten die Ausführungen zum Portmann’schen sekundären Nesthocker gemacht, da hier viele „Quellen“ nur schlechte „Sekundärquellen“ waren, mit Fehlhinweisen, die nur durch mühevolles und eigentlich sinnloses mehrfaches Nachlesen enttarnt werden konnten. Entsprechende Ausführungen habe ich bereits 1981 veröffentlicht [243].
51Innerhalb der Klasse der Säugetiere (Mammalia) kann man drei Unterklassen unterscheiden, dabei werden die Tiere, die ihre Jungen in einer Gebärmutter sich entwickeln lassen und über eine echte Plazenta ernähren, die Eutheria, von denen, die Eier legen (Prototheria mit der einzigen Ordnung Monotremata) bzw. die Jungen im Beutel tragen (Metatheria, Marsupialia) abgegrenzt.
52Mit der Vorhebung im Text folge ich Portmann [164].
53„Anthropoidea“ ist eine andere Bezeichnung für die Unterordnung Simiae der Primates.