Fast jeder von uns hat schon Reisen gemacht. Dabei treffen wir gemeinhin auf
Neues und Fremdes. Betrachten wir ein beeindruckendes Bauwerk, fragen wir
entweder: „Wer wohnt hier?“ oder „Wer hat hier gewohnt?“ Wir interessieren
uns wirklich, begnügen uns aber in der Regel mit einer kurzen Antwort. Ist
der Erbauer berühmt, wird der Erbauer genannt, ist der Architekt berühmt,
wird der Architekt genannt. Wir fragen meist nicht genauer nach und fühlen
uns sogar bei zu vielen Einzelheiten strapaziert. Wir fühlen uns genervt von
dem langatmigen Reiseführer und wollen weiter, um – so bilden wir uns ein
– noch mehr zu erfahren. Mit dieser Einstellung gehen wir an die meisten
Gegenstände unserer Umgebung heran, der Markenname, den wir zu kennen meinen,
bringt uns dazu, dem Produkt zu vertrauen. Mit dem gleichen oberflächlichen
Interesse treten wir Menschen auch an die Natur heran. Wir fragen, wer hat
sie geschaffen, und begnügen uns mit einer einfachen Antwort. Gott hat sie
geschaffen. In unserer mehrheitlich christlichen Welt bekennen wir sogar diesen
Glauben.1 In
anderen Gesellschaften wird auch ein Gott oder aber verschiedene Götter für die Erschaffung
verantwortlich gemacht. Diejenigen von uns, die in der Schule das Wort Evolution
gehört haben, werden sich mehrheitlich wahrscheinlich nicht mit der Geschichte vom
Schöpfergott2
zufrieden geben, sollten wir doch eigentlich wissen, dass unsere natürliche Umgebung
und wir selber das Ergebnis einer sehr langen stetigen Entwicklung sind. Wir
antworten dennoch auf die Frage, wer hat die Natur geschaffen: Gott hat sie erschaffen,
aber sie ist auch Ergebnis der Evolution. Diese Einsicht – Evolution – verdanken wir
dem englischen Forscher Charles Darwin, der im neunzehnten Jahrhundert nach
Beginn der christlichen Zeitrechnung die Evolutionstheorie aufgestellt hat. Ich bin
sicher, dass die große Mehrheit von uns Schwierigkeiten hat, sich Evolution auch
vorzustellen, sich bewusst zu werden, dass wir Menschen – wie alle Pflanzen
und Tiere – uns auf so etwas wie eine Bakterienzelle zurückführen lassen,
und dass wir – wiederum wie alle Pflanzen und Tiere – momentane zufällig
entstandene Endprodukte einer ungerichteten Entwicklung sind. Wir werden
Schwierigkeiten haben zu glauben, dass wir keineswegs perfekt entwickelt sind,
vielmehr die gesamten Informationen unserer Entwicklung in uns tragen, darunter
auch die Informationen, die eigentlich unwichtig oder aber gefährlich für uns
sind.
Insofern erscheint es mir wichtig, sich bewusst zu machen, was Evolution ist und
letzlich bedeutet. Wichtigste Grundlage ist die Erkenntnis, dass überall in der Natur
mehr Nachkommen produziert werden als unbedingt nötig sind, um den Bestand zu
erhalten. Als Beispiel betrachten wir hier nur das häufigste heimische Säugetier, die
Feldmaus. Die Kopfrumpflänge von Microtus arvalis beträgt 9 - 12 cm. Unter
günstigen Bedingungen kann der erste Wurf bereits im Alter von fünf Wochen erfolgen,
danach werfen die Feldmausmütter in rascher Folge, im Sommer alle drei Wochen 4 - 7
Jungtiere. Diese Angaben machen verständlich, warum die Feldmäuse als
Nahrungskonkurrenten gefürchtet sind, können sie sich doch exponentiell vermehren.
Ich sehe den Leser mit dem Kopf nicken, alle 3 Wochen 4 – 7 Jungtiere, exponentielles
Wachstum, wie schrecklich! Was heißt exponentielles Wachstum wirklich. Zum
besseren Verständnis wollen wir das theoretische Vermehrungspotential der
Feldmäuse exemplarisch betrachten. Wir gehen bei unserer Rechnung von der
Minimalannahme aus, dass eine Maus nur einmal wirft und dass die Jungenzahl alle
fünf Wochen nur vier Junge beträgt. Bei einem Geschlechterverhältnis von
Männchen zu Weibchen von etwa 1:1 können wir im Mittel also 2 Männchen und
zwei Weibchen als Nachkommen eines Ausgangspaares erwarten. Es ist leicht
nachzuvollziehen, dass sich nach jeweils fünf Wochen die Mäusepopulation verdoppelt
hat.3 Nach einem
Jahr hätten wir 210, nach zwei Jahren 220 Mäuse und bereits nach nicht einmal 6,5 Jahren wäre die
Schachbrettsituation4
erreicht, 264 Feldmäuse tummelten sich auf unserer Erde. Wir müssten uns also
vorstellen, dass an jeder auf unserer Erde denkbaren Fläche, ob Wasser oder Land 72
Feldmäuse übereinander gestapelt sind. Wir wissen durch eigene Erfahrung, dass es
diese Mäuseschichten nicht gibt. Verantwortlich hierfür ist das Phänomen, das Charles
Darwin natural selection genannt hat. Natürliche Auslese, was können wir darunter
verstehen? Bleiben wir bei unserem Mäusebeispiel. Mäuse sind Nahrung für zahlreiche
Beutejäger, von denen Katzen und Greifvögel (z. B. der tagaktive Mäusebussard,
nachtaktive Eulen) uns allen bekannt sind. Nimmt die Mäusepopulation zu, haben
diese Beutejäger bessere Chancen, sich selbst zu vermehren, der sogenannte
Predationsdruck5
nimmt zu, was sogar einen negativen Einfluss auf die Populationsentwicklung haben
mag, besonders „unvorsichtige“ Mäuse werden ausgerottet, nur diejenigen, die sich zu
verstecken wissen, haben Chancen, ihr Erbgut an die nächste Generation auch
weiterzugeben. Darüber hinaus werden gerade in unseren Breiten die Mäuse schon
Schwierigkeiten haben, den Winter zu überstehen.
Wir können also sagen, dass in den abertausenden Jahren Mäusegeschichte nur diejenigen
Feldmäuse erfolgreich waren, die über das beschriebene Vermehrungspotential auch
verfügten. Dies gilt nicht nur für Mäuse.
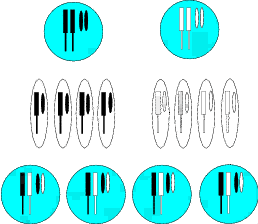
In der Abbildung 2.1 (und den folgenden Abbildungen) ist dies jeweils skizzenhaft
angegeben. Die oberen beiden Kreise symbolisieren die Elterngeneration. Die Hälfte
dieser Chromosomen geben die Eltern an ihre Nachkommen weiter. Die Ovale
kennzeichnen die Geschlechtsprodukte, die man sich als Ei– und Samenzelle vorstellen
kann. Insofern besitzen auch die Nachkommen – die unteren vier Kreise –
auch nur den doppelten Chromosomensatz. Daher muss vor der Bildung der
weiblichen Eizellen bzw. vor der Bildung der männlichen Samenzellen eine
Reduktion des Chromosomensatzes erfolgen. Die vorhergehende Teilung des
Kernes wird daher auch Reduktionsteilung genannt. Sie unterscheidet sich von
der „normalen“ Zweiteilung erheblich. Da – wie bei letzterer – auch bei der
Reduktionsteilung das genetische Material verdoppelt wird, ist das Ergebnis der
Reduktionsteilung eine Teilung in vier Teile. Nun muss sichergestellt werden, dass auch
alle vier Teilstücke identische Chromosomen haben. Dies schafft die Zelle durch einen
komplizierten Mechanismus. Zuerst legen sich die jeweils gleichen Chromosomen direkt
nebeneinander, so dass sie nach verschiedenen Seiten auseinander gezogen werden
können. Klappt dieser Vorgang nicht, führt die Reduktionsteilung nicht zum
Erfolg.
Dieser Mechanismus (vgl. Abbildung 2.2) stellt also auch einen Reparaturbetrieb dar,
bei dem alles Kaputte eliminiert wird. Aus der Skizze können wir erkennen, dass auch
die Nachkommen mit einem Gendefekt – obwohl sie Erbinformationen von beiden
Eltern erhalten haben – sehr wohl zu dieser Reduktionsteilung in der Lage sind.
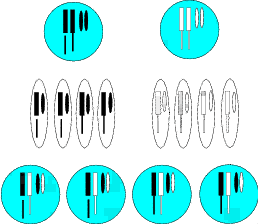
Wir stellen uns nun vor, unsere Art hätte eine Erbinformation „Halte Abstand ..von
dem Wasser.“, diese wäre selektionsbegünstigt, da die Angehörigen mit einer solchen
Information nicht ertrinken (Abbildung 2.2). Bei unserer Inselpopulation wäre diese
Erbinformation aber hinderlich, sie schränkte die Nutzbarkeit der Insel erheblich ein.
Bricht nun bei einem Individuum z. B. ein Chromosom an der Stelle, an der die
Erbinformation „Halte Abstand ...“ steht, dann gibt das Individuum an die Hälfte
seiner Nachkommen drei statt zwei Chromosomen weiter. Die zweite Hälfte der
Nachkommen erhält – wie erwartbar – unverändert zwei intakte Chromosomen
(Abbildung 2.2).
Diesen Vorgang können wir uns sowohl bei Individuen der Festlandspopulation als
auch bei Individuen der Inselpopulation vorstellen. In beiden Populationen hat der
Bruch keine Konsequenzen. Die Reifeteilung der Nachkommen kann genauso
stattfinden, da sich an das ungebrochene Pendant die beiden Teilstücke entsprechend
anlagern können. In beiden Populationen gibt es also Individuen mit vier
Chromosomen, die von beiden Elternteilen jeweils zwei Chromosomen geerbt haben,
und Individuen mit fünf Chromosomen, die von einem Elternteil zwei und von dem
anderen drei Chromosomen mitbekommen haben. An dem genetischen Programm der
jeweils betroffenen Individuen hat diese unterschiedliche Ausstattung nichts geändert,
zwar ist – wie oben ausgeführt – durch den Bruch die Erbinformation „Halte
Abstand....“ verloren gegangen, doch hat dies zu keiner Änderung geführt, da diese
Information auf dem vom anderen Elternteil ererbtem intakten Chromosom
erhalten geblieben ist und dementsprechend weiterhin das Individuum steuert.
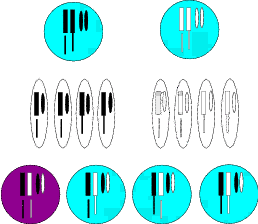
Treffen aber nun in beiden Populationen zwei Tiere aufeinander, die den gleichen
Defekt unsichtbar in sich tragen, dann geben sie jeweils an die Hälfte ihrer
Nachkommen den Defekt weiter. Im Mittel kommt dabei heraus, dass ein Viertel der
Nachkommen überhaupt keinen Defekt mehr hat, die Hälfte ihn unsichtbar mit sich
trägt und ein Viertel die Erbinformation „Halte Abstand vom Wasser“ verloren
hat (Abbildung 2.3). Über die Zeit wird das Fehlen dieser Information für
die Festlandspopulation hinderlich sein, da ein Teil der Nachkommen der
Individuen mit diesem Defekt ertrinken, was deren Fitness reduziert. Für die
Inselpopulation mag das gleiche gelten, einige ertrinken, doch wird dieser
Verlust durch reichliche Nahrung am Inselufer ausgeglichen. Dank der besseren
Ernährung überleben mehr Nachkommen als bei den Individuen, die konsequent
Abstand zum Wasser halten. Die Defektträger sind also fitter. Zwangsläufig
setzen sie sich über die Zeit durch. Unsere ursprüngliche Art wäre also in zwei
Unterarten aufgespalten. Nennen wir die Festlandspopulation „A“ und die
Inselpopulation „B“. In unserem Beispiel unterscheiden sie sich nur in einem einzigen
Merkmal, in der „echten Natur“ wären per Zufall sicherlich weitere Merkmale
betroffen. Wir haben schon ausgeführt, dass nicht alle Arten mit dem Inselleben
zurechtkommen, insofern wollen wir hypothetisch annehmen, auf der Insel
stürben die Konkurrenten unserer Art, die auf den Bäumen lebten, aus. Wir
haben also bei unserer Inselpopulation eine Unterart mit sechs Chromosomen
konstruiert.18
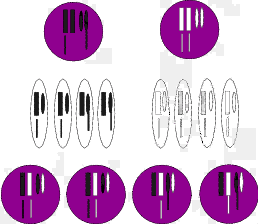
Gehen wir innerhalb der Unterart „B“ nun wieder von einer zufälligen Mutante aus,
bei einem Individuum klebe eines der kleinen Chromosomenstücke an dem zweiten (in
unserer Skizze dem spindelförmigen) Chromosom fest. Die Hälfte der Nachkommen
dieses Individuums hat nun fünf Chromosomen (statt sechs). (Abbildung 2.4) Durch
das Verkleben sei durch Zufall die Information „Kletter auf Bäume“ entstanden, die
die Hälfte der Nachkommen unseres Individuums in sich tragen. Auch hier gibt es bei
der Reduktionsteilung keine Probleme, da sich das bei der beschriebenen
Unterartenbildung entstandene kleine Chromosom sich weiterhin anlagern kann. Die
neu entstandene Mutante wird sich also in unserer Inselpopulation weiter
vermehren. Diese Verhältnisse sind wieder in einer Skizze dargestellt (Abbildung
2.4).
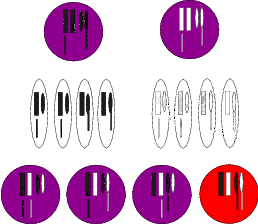
Auch hier wird der Fall eintreten, dass zwei Nachkommen, die die Information „Kletter
auf Bäume“ versteckt mit sich tragen, sich miteinander paaren, das Ergebnis
können wir bereits erwarten, ein Viertel der Nachkommen tragen reinerbig diese
Informationen, haben also nur noch vier Chromosomen. Diese Variante wäre
gleichzeitig eine neue Unterart, die wir als „C“ bezeichnen wollen (Abbildung 2.5).
Wir haben keine Probleme, davon ausgehen zu können, dass die Variante
„C“ besonders erfolgreich sein wird, da sie zusätzlich über weitere Nahrungsoptionen
verfügt.
Auf unserer Insel leben nun zwei Unterarten, die sich nicht in einer Konkurrenzsituation
befinden, Individuen der Unterart „B“ nutzen den Boden, Individuen der
Unterart „C“ die Bäume, sie leben in unterschiedliche ökologischen Nischen.
Für diesen Prozess der Unterartenbildung, der auch tatsächlich, wie wir
noch zeigen werden, für viele Verteter der Primates belegbar ist, benötigt
die Evolution Zeit. Für die auf wenigen Seiten beschriebenen Prozesse
müssen Zeiträume von 100 000 Jahren bis 1 Million Jahre veranschlagt
werden.19
Ein Forscher in seinem Forscherleben wird also nicht die Chance haben, den gesamten
Prozess zu begleiten. Durch die menschlich bedingte Fragmentierung der Regenwälder
ist zu vermuten, dass diese Unterartenbildungen und Artenbildungen beschleunigt
werden, da die Individuen der verschiedenen Fragmente untereinander isoliert sind.
Gleichzeitig haben wir nämlich ein Beispiel für das komplizierte Entstehen von Arten
erhalten. Individuen von Population „A“ und Population „B“ können miteinander
erfolgreich züchten, sind also Unterarten einer gemeinsamen Urart „A“, Nämliches gilt
für Population „B“ und „C“, beide sind Unterarten ihrer gemeinsamen Urart „B“.
Doch treffen Mitglieder der Populationen „A“ und „C“ aufeinander und paaren sich,
dann können ihre Nachkommen keine Reduktionsteilung mehr bestehen, sie sterben
ohne Nachkommen aus. Population „A“ und Population „C“ sind also auf
Artenniveau zu trennen.
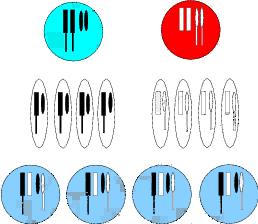
Das merken wir freilich nicht, da Unterart „A“ auf der Insel nicht vorkommt. In der
Erdgeschichte folgen auf Warmzeiten jedoch Eiszeiten, die Wasserbindung an
den Polkappen nimmt zu, der Wasserspiegel sinkt, unsere Insel ist wieder
mit dem Festland verbunden. Nun würde diese Artenbildung offensichtlich
werden.20
Solche Wege der Unterarten- und Artenbildung sind gerade bei den Primates
mehrfach aufzeigbar. Wie bereits besprochen (Kapitel 1) können wir bei den Primaten
zwischen Halbaffen und Echten Affen unterscheiden. Meine ersten Primaten (vgl.
Kapitel 3 und Kapitel 4) waren Halbaffen. Vernachlässigen wir hier die Koboldmakis,
auf die ich schon hingewiesen habe (Kapitel 1), können wir bei den Halbaffen zwischen
den Festlandshalbaffen, den Lorisiformes , und den Halbaffen Madagaskars, den
Lemuriformes, unterscheiden. In der Evolutionsgeschichte der Primaten gab
es auf Madagaskar keine Echten Affen, so dass sich hier die Halbaffen ohne
Konkurrenzdruck durch Echte Affen entwickeln konnten. Unterschiedlichste
Arten eroberten die unterschiedlichsten ökologischen Nischen und nutzten die
unterschiedlichsten Tageszeiten. So leben auf Madagaskar tagaktive neben
nachtaktiven Formen. Sowohl bei den tag- als auch bei den nachtaktiven Formen
werden regelmäßig neue Arten beschrieben, dabei wird in der Regel - dem
angegebenen Beispiel entsprechend - ehemaligen Unterarten nun Artenstatus
zuerkannt.
Die Festlandshalbaffen sind alle nachtaktiv (nocturnal). Offensichtlich konnten auf dem
Festland unter dem Konkurrenzdruck der Echten Affen entweder keine tagaktiven Formen
überleben21
oder sich keine tagaktiven Formen entwickeln. Innerhalb der
Lorisiformes22
dürfen wir wiederum zwischen der Familie Lorisidae und der Familie Galagidae
unterscheiden. Bei den Lorisidae gibt es asiatische (Loris und Nycticebus) und
afrikanische (Perodicticus und Arctocebus) Gattungen, bei den Galagidae nur
afrikanische (Gattung Galago).
Die Lorisidae werden fälschlicherweise auch als Faultiere unter den Primaten
bezeichnet. Diese Bezeichnung ist aber nach meiner Einschätzung falsch. Nach
Beobachtungen an Vertretern aller erwähnten Gattungen möchte ich betonen, dass
grundsätzlich alle lorisiformen Primates sich gleich schnell bewegen können.
Die Galagos haben zusätzlich die Fähigkeit erworben, gut und schnell zu
springen23,
die Loris zusätzlich die Fähigkeit, sich extrem langsam zu bewegen und in der
Bewegung zu verharren. Auf den ersten Blick scheinen diese unterschiedlichen
Spezialisierungen nicht miteinander vereinbar zu sein. Fragen wir aber nach den
Selektionsmechanismen, sind wir uns schnell einig: Es ist das Wichtigste zu
überleben und genügend Nahrung erbeuten zu können. Hier bezeugen die
heute lebenden Lorisiformes zwei unterschiedliche erfolgreiche Strategien. Die
galagoartigen Tiere sind schnelle Springer, können sowohl schnell entkommen als auch
schnell Beute fangen. Die loriartigen stellen bei Gefahr die Bewegungen ein
(und werden übersehbar); treffen sie auf lebende Beute, können sie sich so
langsam annähern und nach der Beute greifen, dass sie von dieser nicht geortet
werden. Im letzten Zugriff sind die Loriartigen dann aber genauso schnell wie
die Galagos. Zu den Galagos gehörten auch meine ersten Primaten. Für das
Anfertigen einer Diplomarbeit erhielt ich Riesengalagos. Riesengalagos sind
etwa kaninchengroß, ihren Namen verdanken sie dem Umstand, dass sie im
Vergleich zu den kleineren Galagos, den typischen Buschbabies, wahre Riesen
sind.
1Glaubensbekenntnis
2Indirekt bekennt schon der seinen Glauben, der von Mitgeschöpfen spricht. Da nichts geschaffen wurde, gibt es auch keine Mitgeschöpfe, man müsste von Mitlebewesen oder Mittieren reden. Viele Theologen haben jedoch ein allzu großes zwanghaftes Bedürfnis, einen Gott für das Leben und Sterben verantwortlich zu machen und diesem dann auch noch zu danken. Ein fast skurilles Beispiel für christlichen Aberglauben hörte ich einmal in einer Rundfunksendung. Der Radiotheologe forderte seine Zuhörer auf, statt über die Autobahn einmal über die Landstraße zu fahren, so könne man die verschiedenen Landschaften besser genießen, die Gott sich ausgedacht habe.
3Wir gehen davon aus, dass die Eltern versterben.
4Das Schachbrett hat 64 Felder, angeblich habe der ägyptische Erfinder des Schachspielens einen Wunsch freigehabt. Er soll sich gewünscht haben, ein Korn Getreide auf das erste Feld, das Doppelte (zwei Körner) auf das zweite Feld, das Doppelte des zweiten Feldes (vier Körner) auf das dritte usw. Die reich gefüllten Kornkammern Ägyptens reichten freilich nicht aus, die Belohnung zu zahlen.
5Der Begriff Predation bzw. auch Predator geht auf das aus dem Lateinischen (praedator) abgeleitete englische Wort predatory = räuberisch zurück, wir kennen vergleichbare Begriffe in unserer Sprache, z. B. „Raubvogel“ oder „Raubtier“, wobei natürlich weder der Vogel noch das Tier tatsächlich rauben.
6Die geringe, aber für uns dennoch verhängnisvolle Nachkommenschaft konnten sich diese Schaben wohl wegen ihrer versteckten Lebensweise leisten, die systematisch nahe verwandte in wärmeren Gebieten freilebende Gottesanbeterin Mantis religiosa benötigt im Mittel mehr als 1000 Nachkommen pro Weibchen, um uns auch heute noch mit ihrer Anwesenheit zu erfreuen.
7Wegen des hohen individuellen Alters der Meeresschildkröten - sie werden älter als wir Menschen - werden wir in unserer Generation ihr schleichendes Aussterben möglicherweise nicht bemerken. Einige Meeresschildkröten müssen nicht nur sehr alt werden und regelmäßig ihre etwa hundert Eier umfassenden Gelege liefern, um sicher zu stellen, dass im Mittel pro Schildkrötenweibchen zwei Junge überleben. Vielmehr müssen und mussten sie dies auch noch zur gleichen Zeit und am gleichen Ort gemeinsam mit tausenden Artgenossen leisten. Nur so erreichen sie, dass eine hinreichende Zahl von Nachkommen selber fortpflanzungsfähig wird. Auf die Schildkrötenbabies warten nämlich schon alle möglichen Beutejäger. Nur bei einer genügend großen Zahl von gleichzeitig aus dem Sandstrand in das Meer drängenden Jungen, haben einige die Chance, zum auch nur relativ sicheren Meer zu gelangen. Über mehr als 100 Millionen Jahre lang haben nur die Schildkrötenweibchen, die genügend viele Eier legten, durch ihre Nachkommen bis heute überlebt. Erst unsere Enkel und Urenkel werden wissen, ob es heute hinreichend große Zahlen von Meeresschildkröten gibt.
8Zu dem Namen „Spanner“ sind die zu dieser Gruppe gehörenden Schmetterlinge wegen der Fortbewegung ihrer Raupen gekommen.
10Es muss sich nicht immer um Lernvorgänge handeln, möglicherweise folgt der Beutejäger vielleicht nur einem festen - via Selektion - entstandenem genetischen Programm.
11Den schnellen Zugriff zum Internet gab es damals noch nicht.
12Zum Nachweis muss man den Baum jedoch fällen. Das Besteigen von morschen Bäumen ist kaum ratsam.
13Hier verdanken wir wichtige Einsichten wiederum Willi Hennig[74]: „A characteristic of any science ist the ‚endless task‘ and the knowledge that its final goal will probably never be reached. This is also true of phylogenetic systematics.“ Seite 28[74].„Thus by definition phylogenetic relationships exist only between species; they arise through the process of species cleavage. The key position of the species are, in the sense of the class theory, the elements of the phylogenetic system.“Seite 29 [74].
14Im Gegensatz zu Darwin müssen wir uns nicht mit der Feststellung, dass Variabilitäten auftreten, zufrieden geben.
15Die Einschränkung „in der Regel“ ist notwendig, da manchmal Männchen und Weibchen sich im Aussehen erheblich unterscheiden, so dass anfänglich beide Geschlechter als verschiedene Arten beschrieben werden.
16Neben den Erbinformationen des Zellkerns gibt es noch solche in der Eizelle, insofern sind wir auch näher mit unserer Mutter verwandt als mit unserem Vater, von dem wir nur die Informationen des Zellkerns erben.
17Bei Pflanzen können mehrfache Chromosomensätze vorhanden sein
18In einem Schulbuch würde aus didaktischen Gründen gezeigt, dass bei sechzehn Nachkommen das Verhältnis 4 : 8 : 4 wäre.
19Evolution hat also Zeit. Dies erklärt auch, warum bei unseren Haustieren und deren relativ kurzer Domestikationsgeschichte noch keine neuen Arten entstanden sind. Dabei kann natürlich nicht ausgeschlossen werden, dass die eine oder andere Rasse eventuell unbemerkt bereits den Übergang zu einer neuen Art beschritten hat.
20Als Betrachter der etwa 70 Millionen Jahre andauernden Primatenevolution fragt man sich schon, warum sich die Politik gedanklich um die Erwärmung der Atmosphäre sorgt. Nach meinem Wissen steuern wir tatsächlich – unbeeinflussbar von uns Menschen – auf die nächste Eiszeit zu. Dieses Zusteuern dauert freilich. Es ist in einem Menschenleben fast nicht bemerkbar. Vielleicht aber wird der Beginn dieses Prozesses auch durch die menschlichen Emissionen maskiert.
21Bei Fossilfunden ist es nicht möglich, Aussagen über die Aktivitätszeiten zu leisten.
22Namensgebend ist hier der Schlanklori Loris tardigradus.
23Zum Springen sind sie besonders durch die Ausbildung eines „Sprungbeins“ befähigt, zwei Mittelfußknochen (Naviculare und Calcaneus) sind auffällig verlängert.